






1. INTRODUCTION
Mushrooms are widely regarded as a superfood, serving not only as a nutritious and protein-rich dietary source but also as an important resource for pharmaceutical applications (Ambhore et al., 2024; Magaña Amaya and Shimizu, 2025). They play a vital role in rural communities, particularly for mushroom farmers, because of the significant economic benefits derived from their cultivation and trade.
The cultivation of edible and medicinal mushrooms on lignocellulosic materials is an important biotechnological process that contributes to sustainable waste management and high-value biomass utilization. Sawdust from various wood species is among the most widely used substrates for mushroom cultivation because of its suitable texture, high porosity, and availability as a byproduct in the wood industry. Species, such as Auricularia spp. (commonly known as wood ear mushrooms) and Ganoderma lucidum (lingzhi or reishi mushrooms), are well known for their ability to colonize woody substrates and for their pharmacological properties, including antioxidant, immunomodulatory, and anti-inflammatory effects.
Indonesia is a tropical country with abundant forest resources and a well-developed wood-based industry. The country’s forests are rich in biodiversity, including various wood species with high extractive content, which is crucial for multiple industrial applications. Many Indonesian wood species are known for their high extractive content, which contributes to their natural durability and aesthetic quality (Arisandi et al., 2025; Fernandes et al., 2024; Iswanto et al., 2021). Sawmilling generates substantial quantities of low-value wood residues. Through the fungal degradation of lignocellulosic biomass, these residues can be converted into value-added products (Gwak et al., 2022, 2024; Sneha and Tomy, 2017; Sutapa et al., 2024; Zakaria et al., 2022). The utilization of forest industry byproducts in specialty mushroom cultivation effectively integrates forestry and agricultural systems and supports the principles of a circular economy (Grimm and Wösten, 2018). As a lignocellulosic material rich in cellulose, hemicellulose, and lignin, wood is an excellent carbon source for fungal growth. The use of sawdust residues not only supports mushroom production but also contributes to environmental pollution mitigation (El-Ramady et al., 2022).
The successful growth of fungal mycelia on sawdust-based media largely depends on the physicochemical characteristics of the wood, particularly its lignocellulosic composition and secondary metabolites. Different mushroom species prefer different substrates. For example, medicinal mushrooms, such as shiitake and oyster mushrooms (pearl and yellow varieties), grow better on straw and other agricultural residues, whereas reishi and maitake grow better on sawdust and logs. Conversely, Agaricus mushrooms grow on manure, similar to button mushrooms and portobello mushrooms (Olaniran et al., 2024). In addition, mushrooms grown on substrates rich in hemicellulose typically have low fat and high crude fiber contents (Im and Han, 2023; Rawiningtyas et al., 2023).
Among the chemical components of wood, wood extractives—a diverse group of low-molecular-weight organic compounds, including phenolics, tannins, resins, fats, and terpenoids—play a critical yet often overlooked role. These compounds vary greatly among wood species and can either stimulate or inhibit fungal growth; n-hexane-soluble extracts from teak inhibit mycelial growth (Lukmandaru, 2015). This is consistent with the observation that mushroom farmers often avoid certain local wood residues as substrates, likely because of their inhibitory extractive compounds, such as mahogany, teak, jackfruit, mango, and black rosewood. However, sawdust from these species is present in considerable quantities. Nevertheless, not all extractives are detrimental; some have been reported to stimulate the mycelial growth of Pleurotus florida (Atila et al., 2024). Conversely, enrichment with medicinal plants, such as lemongrass, neem, citrus lemon, and eucalyptus, at levels of up to 4% for each component in the substrate compost used for the cultivation of Pleurotus ostreatus and P. florida, revealed that such addition not only enhanced the yield but also reduced the incidence of contaminants (Aparna et al., 2022). Certain phenolic extracts may exhibit antimicrobial or antifungal activity, thereby suppressing mycelial development, whereas others may serve as additional carbon or energy sources that enhance fungal metabolism (Suprianto et al., 2023; Yoon and Kim, 2023).
Although several studies have investigated the effects of lignin, cellulose, and hemicellulose in mushroom cultivation, research focusing specifically on wood extractives and their direct effects on mycelial growth remains limited. This gap in understanding is particularly relevant in tropical regions, where hardwoods typically contain higher levels of extractives that may influence mycelial growth. Moreover, the assessment of mycelial growth on extracted wood using hot water and ethanol–toluene extraction provides a controlled approach to evaluate the biological response of fungi without interference from environmental variables. This study aimed to evaluate the influence of wood extractives derived from different wood species on the mycelial growth of Auricularia spp. and Ganoderma lucidum. The findings are expected to help determine the best combination of wood meal pre-extraction methods and wood species suitable for the cultivation of each mushroom species. These findings are further supported by the potential inhibitory or stimulatory effects of specific extractive components on fungal development. This knowledge will contribute to the optimization of substrate formulations and the improvement of the efficiency and sustainability of mushroom production systems.
2. MATERIALS and METHODS
Wood residues in the form of sawdust from mango (Mangifera indica) = W1, mahogany (Swietenia mahagoni) = W2, black rosewood (Dalbergia latifolia Roxb.) = W3, and jackfruit (Artocarpus heterophyllus) = W4 were collected from wood industries in the Bantul and Sleman districts, respectively. Spawns of wood ear mushrooms (Auricularia spp.; M1) and lingzhi or reishi (Ganoderma spp.; M2) were supplied by a local supplier (PT Volva Indonesia) in Yogyakarta, Indonesia. Specific strain information was not available. Identification was based on macroscopic morphological characteristics, including pileus morphology, coloration, and texture, following standard taxonomic keys (Kirk et al., 2008). Molecular identification was not performed; therefore, the samples were designated at the genus level.
To remove both polar and nonpolar compounds, 10 g of sawdust from each species was subjected to Soxhlet extraction with 250 mL of ethanol–toluene (1:2) for 6 h (4–6 cycles per hour) until it was free of extractives, based on ASTM standard D1107-96 (ASTM, 2007).
For the removal of water-soluble compounds, 10 g of sawdust was placed in a 300 mL Erlenmeyer flask with 50 mL distilled water and then heated in a water bath equipped with a reflux condenser for 3 h until the extractives were eliminated.
Sawdust was ground to 40–80 mesh and analyzed for ethanol–toluene soluble extractives, hot-water soluble extractives, holocellulose, α-cellulose (ASTM, 1984), and Klason lignin (TAPPI 222 om-02). Extracts from the ethanol–toluene fraction were further evaporated using a rotary evaporator at 60°C. The hot-water solution was evaporated on a hot plate until the solvent had evaporated. The extract was dried in an oven (103°C ± 2°C), and the extractive content was quantified and analyzed using gas chromatography–mass spectrometry (GC–MS; Shimadzu QP2010, Shimadzu, Kyoto, Japan). The samples (1 μL) were derivatized before injection, i.e., 100 μL of pyridine and 100 μL of N,O-bis(trimethylsilyl)trifluoroacetamide with 1% trimethylchlorosilane were combined with 2 mg of each dried ethanol–toluene and hot-water extract. The resulting mixtures were then heated to 103°C for 30 min and ultrasonicated at room temperature. Measurement conditions: RTx-5 capillary column (30 m × 0.25 mm I.D., 0.25 μm); column temperature 70°C; injection temperature 290°C; temperature programming: initial temperature 50°C (held for 5 min) to final temperature 300°C at a rate of 5°C/min (held for 12 min); interface temperature 270°C; split ratio 1:0; pressure 108.1 kPa; helium gas was used as the carrier gas. Component identification was based on the National Institute of Standards and Technology database library (National Institute of Standards and Technology 11 library version). Quantification of each component was based on the percentage area of each GC–MS peak. Ash content and pH values were also determined.
Media for M1 and M2 were formulated from wood meal of each of the four wood species, rice bran (12.5% w/w of oven-dry sawdust weight), calcium carbonate (6% w/w), and water to adjust the moisture content to 70%. Each wood species was subjected to extraction using hot water and ethanol–toluene to obtain hot-water-extractive-free wood meal (treatment 1, T1) and ethanol–toluene-extractive-free wood meal (treatment 2, T2), respectively. The control consisted of a growth medium prepared from untreated wood meal that retained its natural extractives. Portions of 20 g were placed into 90 mm Petri dishes, compacted, and a central cavity was made for inoculum placement. The media were sterilized by autoclaving at 121°C and 1 atm for 15 min. Each treatment was replicated five times.
Sterilized media were inoculated with mushroom spawn and incubated in the dark at 25°C. Colony diameter was measured every 2 d in four directions until complete colonization of the Petri dish (Irawati et al., 2012).
The glucosamine content was used as an indicator of mycelial biomass. This approach is based on the presence of glucosamine in the chitin of fungal cell walls, which is correlated with the amount of fungal biomass (Chysirichote et al., 2014). After 50 d of incubation, 1 g of dried medium was hydrolyzed with 5 mL of 5 M HCl at 80°C for 20 h to degrade chitin in fungal cell walls into N-acetylglucosamine. The concentration was quantified using the braid-and-line method (Ohga, 2000).
For chemical composition analysis, a completely randomized design with a single factor (wood species) was applied, and the data were subjected to one-way analysis of variance using SPSS 16.0. For mycelial growth analysis, a factorial completely randomized design was employed with two factors: mushroom species and wood species. The chemical component data from the GC–MS analysis are presented descriptively.
3. RESULTS and DISCUSSION
Four wood species commonly used in the Indonesian wood industry were selected. Mango and jackfruit wood originate from fruit trees that are widely cultivated by local communities in home gardens and private plantations. These species are characterized by their distinct coloration—light pink for mango and yellow for jackfruit—and are typically used to construct window frames, doors, and other household components. They are preferred because of their characteristic color and high natural durability (Baliga et al., 2011; Sami et al., 2025). Mahogany and black rosewood are also extensively planted by communities as well as in private and state-managed forests. Both are widely employed in the wood industry for construction, furniture manufacturing, and other applications because they are easy to process and have excellent natural durability (Arunkumar et al., 2022; Yanti et al., 2019). Although these species produce large quantities of sawdust as an industrial byproduct, this material is rarely used by mushroom growers as a cultivation substrate because of its slow mycelial growth. Several wood species with excessive levels of extractives, particularly antifungal compounds, may suppress mycelial growth by creating a substrate environment that is not conducive to fungal development (Bopenga Bopenga et al., 2020; Brocco et al., 2025; Jung et al., 2025).
The chemical composition of the residues from the four wood species—mango, mahogany, black rosewood, and jackfruit—used in this study is presented in Table 1, along with the composition of rice bran, which served as a nutritional supplement in the cultivation medium. Statistically, only the acid-soluble lignin content showed no significant differences among the four species.
The chemical profiles of the four wood species (W1–W4) showed statistically significant differences across all parameters. Hot-water extractives were significantly higher in W1 and W4 (11.2 ± 0.7–11.5 ± 0.3%). Ethanol–toluene extractives were highest in W3 (10.0 ± 0.3%). For structural carbohydrates, W3 exhibited the highest holocellulose and α-cellulose contents (70.0 ± 0.2% and 44.3 ± 0.3%, respectively), indicating a superior polysaccharide composition, whereas W1 and W2 formed the lowest statistical groups. Hemicellulose showed an opposite pattern: W3 had the lowest value (25.7 ± 0.1%), whereas W2 (31.9 ± 0.2%) had the highest among the wood species. Lignin content also differed markedly, with W2 (27.1 ± 0.5%) forming the highest statistical group. Acid-soluble lignin exhibited no meaningful statistical trend owing to its very low variation. Ash content varied significantly, with W2 (13.7 ± 0.2%) showing the highest values. pH values clustered into three groups: slightly acidic in W1 (5.5 ± 0.03), moderately acidic in W3 (6.4 ± 0.02), and neutral in W2 and W4 (7.5 ± 0.02). Rice bran consistently differed from all wood groups, with the highest holocellulose, hemicellulose, lignin, and ash contents.
The chemical composition of the wood species analyzed in this study was generally consistent with that in previous reports. The ethanol–toluene extractive and holocellulose contents of mango wood were comparable to those reported by García-Mahecha et al. (2023). The lignin content of mahogany wood was lower than that of African mahogany (Santos et al., 2020). Similarly, black rosewood exhibited lower holocellulose, α-cellulose, and lignin contents than those reported for Dalbergia oliveri (Wei et al., 2022). Jackfruit wood showed lower ethanol–toluene extractive, holocellulose, and α-cellulose contents but higher hot-water-soluble extractive and lignin contents than those reported by Chen et al. (2011).
Wood-decaying fungi typically utilize cellulose and lignin as carbon sources. Wood extractives may exert either positive (including soluble carbohydrates, vitamins, and organic salts) or inhibitory (including polyphenols, terpenoids, tropolones, and tannins) effects on fungal growth (Molokoane et al., 2023). In addition to the proportion of extractives present in wood, the chemical nature of these compounds influences mushroom mycelial growth. Therefore, GC–MS analysis was conducted to identify the chemical constituents of each extract.
The GC–MS analysis of the hot-water and ethanol–toluene extracts from the four species is summarized in Tables 2 and 3. Table 2 shows that the GC–MS chromatogram of W1 was dominated by 4-hydroxy-4-methyl-2-pentanone (33.0%), indicating a high abundance of hydroxy ketone derivatives. Carbohydrate-related compounds, such as D-glucose derivatives (12.6%) and 4,6-benzylidene-α-methyl-D-glucoside (14.1%), also appeared in substantial amounts, suggesting partial depolymerization of wood polysaccharides. Aromatic ester derivatives, including benzoic acid thiazolidinyl ester (12.7%), contributed to the remaining profile. No dominant phenolic or resin acids were observed in W1. The chromatogram of W2 showed a strong presence of 4-hydroxy-4-methyl-2-pentanone (28.1%). Several small polar compounds, such as 1,3-dioxolane (20.6%), isopentanal (10.1%), and 1,6-dideoxydulcitol (10.1%), were identified as sugar degradation products and low-molecular-weight volatiles. Trimethylsilylated silicic esters and silylated benzoic acid derivatives (approximately 10%) suggested that inorganic–organic complexes were formed during derivatization. W3 contained the highest proportion of 4-hydroxy-4-methyl-2-pentanone among all samples (34.1%). Notably, W3 also exhibited very high peaks for 1,4,7-androstatrien-3,17-dione (37.9%), a steroid-like compound, and 2,4,6-trimethyl-1,3-benzenedimethanol (23.6%). Minor amounts of β-hydroxy-isovaleric acid and silylated hydroxyphenyllactic acid (~2%) were present. The dominance of steroidal and aromatic diol compounds distinguished W3 from the other wood species. The chromatogram of W4 displayed a wide distribution of compounds, each present at 7.4%–7.9%, with no single dominant peak. Its constituents included anthraquinone derivatives, protocatechuic acid, diphenylpropenone, hydroxymandelic acid, and steroidal hydrocarbons. Many of these compounds are polyaromatic quinones and phenolics traditionally associated with hardwood extractives. The presence of multiple quinone compounds suggests high oxidative and antimicrobial potential.
Table 3 shows that the ethanol–toluene extractives of W1 displayed a highly diverse chemical composition characterized by numerous compounds with relatively balanced peak intensities. The major constituents included benzene derivatives, such as 1,3-dimethylbenzene (16.74%) and benzaldehyde (15.87%), as well as the aliphatic hydrocarbon nonane (14.27%). Several oxygenated aromatic and phenolic derivatives were also detected, including benzyl alcohol (9.58%) and multiple trimethylsilyl esters of benzoic acid. The presence of both aromatic hydrocarbons and oxidized phenolics indicates that W1 contains a mixture of lignin-derived volatiles and partially oxidized aromatic compounds. Aliphatic hydrocarbons (octane isomers and dodecane derivatives) and siloxanes were observed in moderate proportions. W2 was dominated by a smaller group of compounds with highly skewed relative abundances. The most abundant constituents were p-xylene (65.15%), o-xylene (27.11%), and 2(R),3(S)-1,2,3,4-butanetetrol (26.45%), resulting in a profile dominated by xylene isomers and sugar alcohol derivatives. Other benzoic acid trimethylsilyl esters were also present at lower concentrations.
The prevalence of xylenes suggested a strong contribution from lignin-derived aromatic fragments, whereas sugar alcohols reflected holocellulose-derived degradation products. Minor peaks included siloxanes and structurally complex aromatic steroids, but their contributions were <1%. W3 exhibited a very simple chemical profile dominated almost entirely by a single compound, 2,6-dimethoxyphenyl-β-phenylpropionate (89.94%), with only one additional compound present at notable levels, 1-(2-methylpropoxy)-2-propanol (10.06%). This dominance suggests highly specific extractive chemistry, likely originating from specialized phenolic esters. Compared with the other species, W3 contained far fewer GC–MS-detectable components. W4 showed a more complex profile, similar to that of W1 but with a different distribution pattern. The dominant compounds were p-xylene (64.9%) and o-xylene (26.5%), which together accounted for more than 90% of the total peak area. Several benzoic acid trimethylsilyl esters and siloxane derivatives were also detected in the range of 19% to 22%, although these appeared to represent overlapping or repeated peaks of similar chemical identities within the dataset. Minor peaks included ethylbenzene, cumene, and other aromatic or siloxane derivatives. This profile indicated that the W4 extractives were strongly dominated by simple alkyl benzenes and aromatic esters.
Extractives can be divided into lipophilic and hydrophilic fractions, although their boundaries are often indistinct. Lipophilic compounds include fats, waxes, terpenes, terpenoids, and long-chain aliphatic alcohols, which are typically isolated using nonpolar solvents, such as diethyl ether or dichloromethane. Hydrophilic compounds comprise phenolics (tannins, lignans, stilbenes), soluble carbohydrates, proteins, vitamins, and inorganic salts (Candelier et al., 2020).
Mycelial growth was evaluated using 20 g of the cultivation medium in Petri dishes. Measurements were performed using a digital caliper every 2 d along four radial directions. The observations were terminated when the mycelial front reached the edge of the Petri dish. The mycelial growth rates of M1 and M2 are shown in Table 4.
The average mycelial growth rate of M1 ranged from 0.91 to 7.41 mm day–1. The highest mycelial growth was observed on substrates derived from W1T1, W1T2, W3T2, W4T1, and W4T2. Mycelial growth on pretreated substrates was 2.08–6.94 times faster than that on untreated substrates. The highest enhancement was observed for W2T1, indicating that untreated W2C was the least suitable substrate for M1, whereas hot-water extraction markedly improved its suitability.
Similarly, the mycelial growth rate of strain M2 ranged from 0.79 to 7.40 mm day–1. The highest mycelial growth rates were observed in W1T1, W1T2, W2T1, and W3T2. Growth enhancement ranged from 1.64 to 9.42 times compared with that on untreated substrates, with the highest increase observed in W3T2. These findings indicate that W3 was initially unsuitable for M2 cultivation but became favorable after ethanol–toluene extraction.
Mycelial growth rate represents the speed of hyphal extension and is influenced by factors, such as substrate rigidity and environmental conditions. This parameter is commonly used to assess growth inhibition in culture media. Research on Shorea sp. wood reported a mycelial growth rate of 3.33 mm day–1 for A. polytricha (Irawati et al., 2012), which is higher than that observed on untreated substrates in the present study but lower than that on extracted substrates. This enhancement is likely attributable to the removal of inhibitory extractive compounds (Schoder et al., 2024).
No statistically significant differences were observed between the two mushroom species used in this study. However, within the same mushroom species, both wood species and extraction treatments had pronounced effects on mycelial growth. Statistical analysis revealed significant effects of wood species, pretreatment method, and their interaction at the 1% significance level. This indicates that the most appropriate extraction treatment may differ depending on the wood species used. Post hoc Tukey’s honestly significant difference (HSD) analysis identified specific combinations of wood species and extraction methods that differed significantly, as illustrated in Fig. 1.
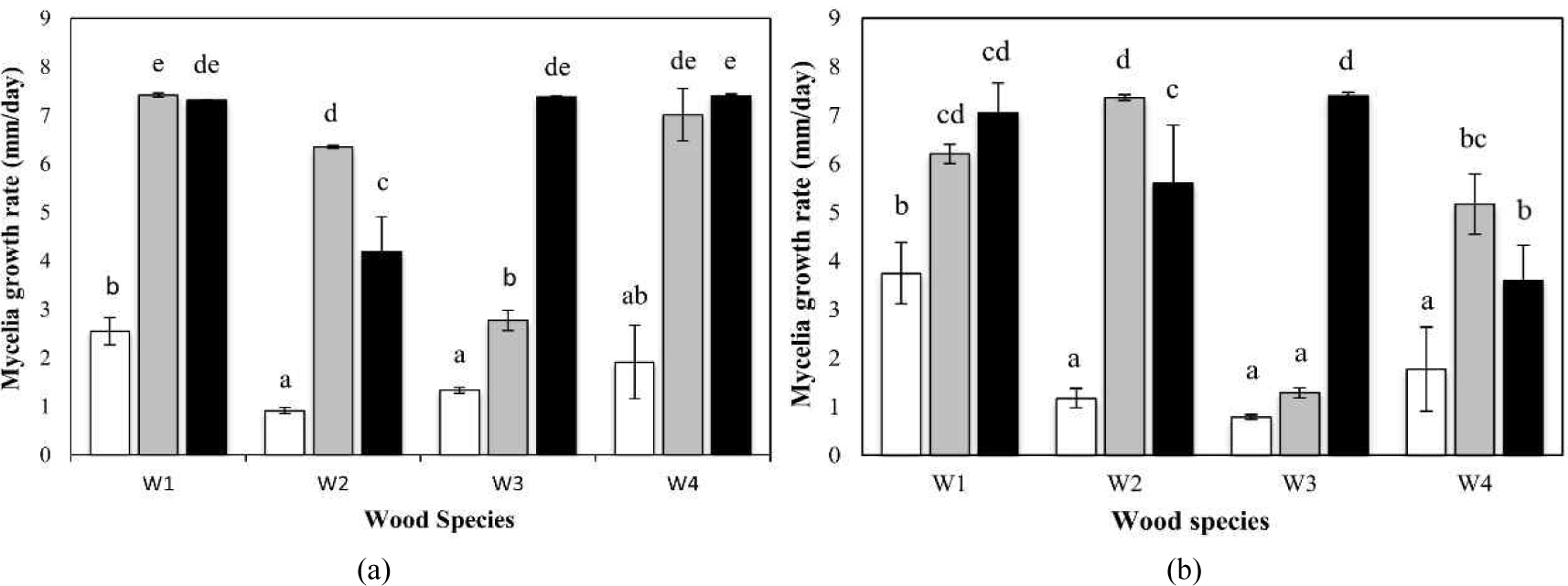
For W1 and W4, comparable improvements in mycelial growth were observed in substrates extracted with either hot water or ethanol–toluene, suggesting that the extractives removed by both treatments exerted similar effects on fungal growth. By contrast, for W2, higher mycelial growth rates were recorded for both M1 and M2 on hot-water-extracted substrates. This result indicates that hot-water-soluble extractives, particularly 4-hydroxy-4-methyl-2-pentanone (Table 2), strongly inhibit mycelial development. 4-hydroxy-4-methyl-2-pentanone may inhibit mycelial growth by inducing solvent stress and altering membrane permeability, thereby disrupting fungal metabolism. Although it is not a strong antifungal compound, its presence may enhance inhibitory effects when acting synergistically with other wood extractives (Cowan, 1999). The opposite trend was observed for W3, where substrates prepared from ethanol–toluene-extracted wood showed more rapid mycelial growth than those extracted with hot water. This suggests that ethanol–toluene-soluble extractives in W3, notably 2,6-dimethoxyphenyl-β-phenylpropionate (Table 3), exert inhibitory effects on the mycelial growth of both M1 and M2. This observation agrees with previous reports that identified toxic extractives in Dalbergia latifolia that inhibit fungal activity (Masendra et al., 2020).
The glucosamine concentration serves as a proxy for estimating fungal biomass (Ohga, 2000). The quantity of mycelia was estimated by measuring the glucosamine content. The glucosamine content in samples incubated for 50 d was measured to evaluate mycelial quantity immediately before flushing. A high mycelial biomass indicates a dense, robust mycelial network, which may develop at different rates depending on the conditions.
The average glucosamine content of M1 ranged from 40.4 to 866.9 μg g–1 (Table 5). The highest absolute glucosamine concentration was observed in W2T1 samples. However, the greatest relative increase in glucosamine content (13.12-fold) occurred in W3T2 compared with W3C. Hot-water extraction enhanced glucosamine levels in substrates derived from W2 and W4, whereas for W1 and W3, ethanol–toluene extraction resulted in greater increases than hot-water treatment.
For M2, glucosamine content ranged from 36.6 to 786.4 μg g–1. In contrast to M1, the highest glucosamine concentration in M2 was recorded on substrates prepared from W3T2. The largest increase in glucosamine content was also observed in W3T2, reaching a 21.48-fold increase. Substrates derived from W3 and W4 that were subjected to either hot water or ethanol–toluene extraction were generally suitable for M2 growth. These findings indicate that the most effective extraction treatment depends not only on the wood species but also on the mushroom species and therefore cannot be generalized across substrates.
A high mycelial growth rate does not necessarily correspond to high glucosamine content (Magaña Amaya and Shimizu, 2025). In addition to identifying growth inhibition factors, this analysis reflects the nutritional suitability of the cultivation medium. Rapid radial expansion may result in a sparse hyphal network and low biomass, whereas slower growth may produce a dense and compact mycelial mat with higher biomass accumulation. Studies on Falcataria molucana wood substrates reported relatively slow mycelial growth of A. polytricha accompanied by high glucosamine content (860 μg g–1; Irawati et al., 2012), a value comparable to that observed for M1 grown on W2T1 in the present study.
No statistically significant differences were detected between the two mushroom species examined. However, within each mushroom species, both wood species and extraction treatments exerted a significant influence on glucosamine content. Statistical analysis revealed significant effects of wood species, pretreatment method, and their interaction at the 1% significance level. This indicates that the most suitable extraction treatment may vary depending on the wood species used. Post hoc Tukey’s HSD analysis identified specific combinations of wood species and extraction treatments that differed significantly, as illustrated in Fig. 2.
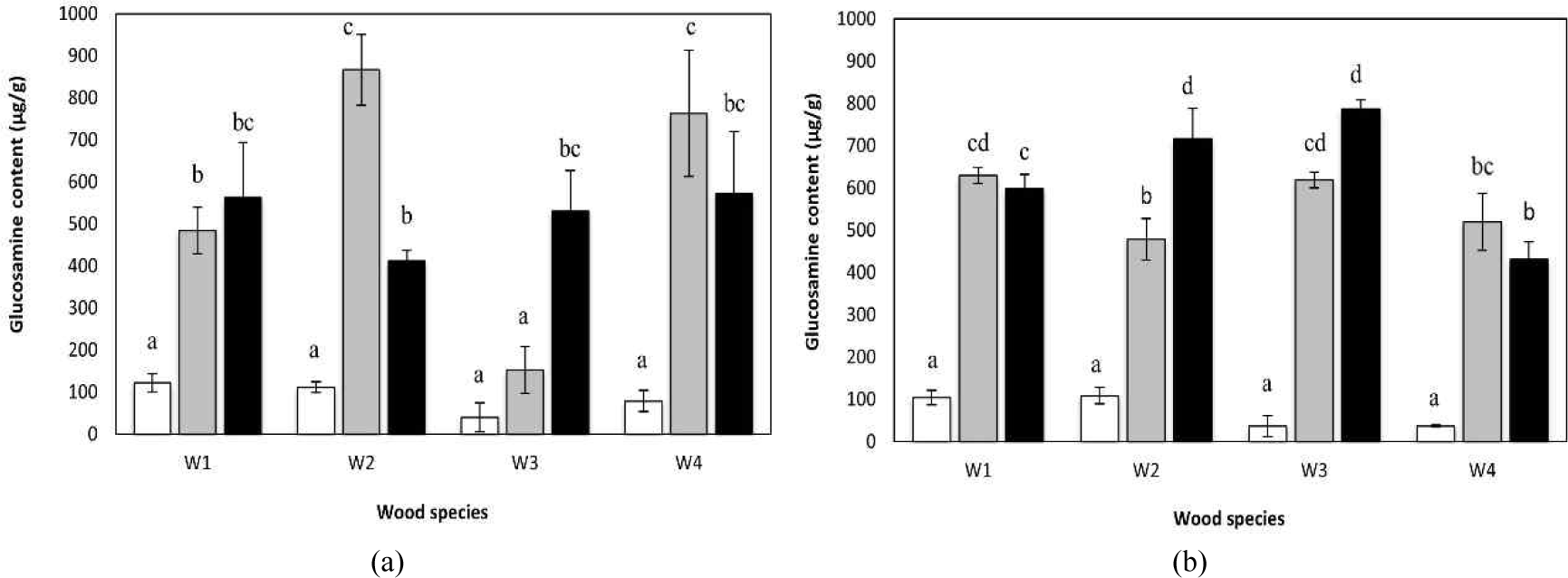
Fig. 2 shows that suitable substrates for M1 growth include W1T2, W2T1, W3T2, and W4T1. By contrast, the optimal substrates for M2 growth were W1T1, W2T2, W3T1, or W3T2.
4. CONCLUSIONS
Wood extraction treatments significantly improved the suitability of wood residues as substrates for mushroom cultivation. The maximum mycelial growth rates of M1 and M2 reached 7.41 and 7.40 mm day–1, respectively, while the highest glucosamine contents were 866.9 and 786.4 μg g–1, respectively. Mycelial growth of M1 increased by up to 6.94-fold in W2 following hot-water extraction, whereas M2 growth increased by up to 9.42-fold in W3 after ethanol–toluene extraction. Similarly, glucosamine content increased by 13.12-fold and 21.48-fold for M1 and M2, respectively, in ethanol–toluene-extracted W3. The compound 2,6-dimethoxyphenyl-β-phenylpropionate present in W3 extractives was identified as a major growth-inhibitory component.