






1. INTRODUCTION
Termites are the most important invertebrate in the decomposition of wood, global carbon cycling, and nutrient recycling and have extraordinary ecological impacts (Bignell et al., 2011; Khan and Ahmad, 2018). Termites’ ability to metabolize cellulosic materials, on the other hand, implies that they can become pests when they attack structural or wooden parts in buildings and other facilities vital to humans. Currently, 3106 living and fossil termite species have been identified (Krishna et al., 2013). More than 3,000 termite species have been identified in the tropics, subtropics, and temperate regions (Su and Scheffrahn, 2000). However, only 183 species substantially damage buildings, furniture, and other wooden materials (Hassan and Morrell, 2021).
Based on their habitat, termites can be classified into three general categories: Subterranean, damp-wood, and dry-wood termites (Baker and Marchosky, 2005). Subterranean termites are closely associated with the soil habitat where they tunnel to locate water and food, such as wood, fallen logs, and other cellulose-containing materials (Suiter et al., 2009) and also have a habit of moving by forming networks in the ground (Im and Han, 2020). Besides being found in nature, termites are also found in human settlements, where termite attacks various valuable properties. Therefore, subterranean termite species are well-known as the most destructive and economically important pests of wood and other cellulose products (Adfa et al., 2020; Peterson et al., 2006).
Most species of termites forage cellulose material. Consequently, baiting termites with cellulose-rich dead wood or cardboard is a successful method for sampling and monitoring termites because of their foraging behavior (Davies et al., 2013; Galbiati et al., 2005). Baiting is also effective for monitoring termite activity over time and is successful in various habitats (Davies et al., 2021). Furthermore, the palatability of the baiting material used is an important factor affecting bait effectiveness. Woods are the most common baiting material for termites and could be economically feasible materials for termite baiting. However, woods have different characteristics, which can influence termite preference. Termite resistance to woods was affected by wood species, and some termite species favor certain wood species over others (Febrianto et al., 2015; Morales-Ramos and Rojas, 2001). Pine is the most frequent type of wood used for baiting. Pine, for example, has been used as commercial termite bait in Sentricon Colony Elimination System® and Sentri Tech® products (Jones et al., 2005). In addition, the research of Arinana et al. (2021) also used pine wood as an ingredient for the formulation of termite pellet bait. However, wooden baits are typically made from wood species known to be susceptible to termites and are available in the location (Jones et al., 2005). Furthermore, identifying local species that have potential as wood bait is required to diversify baiting material.
Studies on termite preferences for local wood species have been carried out by several researchers. The alternative wood species are the potential to substitute pine as wood bait are species of wood originating from community forests. In addition, most species of wood from community forests in Indonesia have low durability and are classified as Durability Class IV–V (Arinana et al., 2012; Jasni and Rulliaty, 2015). Community timbers can be an alternative wood bait to replace pine because they are a fast-growing wood species (Martawijaya et al., 2005). In addition, logs production from community timber has increased from year to year. It was recorded that in 2013 the production of logs from community forests was 28,398,137 m3, and in 2017 it was 37,798,711 m3 (Ministry of Environment and Forestry, 2018). In Indonesia, community forests provide the timber sector with significant wood supplies for industry, such as sawn timber and furniture (Maryudi et al., 2017). Some widely used community timber species are Durio zybetinus, H. brasiliensis, Altingia excelsa, Toona sureni, M. eminii, F. moluccana, Tectona grandis, Swietenia macrophylla, Dalbergia latifolia, P. merkusii and other species (Hakim et al., 2009). Arinana et al. (2012) also used wood species from a community forest in their research and reported that subterranean termites preferred rubberwood (H. brasiliensis), sengon (F. moluccana), and pine (P. merkusii) in laboratory tests.
In addition, the length of the baiting time is an essential factor that needs to be considered. Field testing generally requires three months as the baiting time, such as the research conducted in Bekasi (Arinana et al., 2015), Bogor (Hadi et al., 2016), Bogor (Arinana et al., 2020), and Jakarta (Arinana et al., 2016). Peralta et al. (2004) conducted field tests on Pine sp. and Eucalyptus spp., with the baiting time of 30 days, 45 days, and 60 days. Meanwhile, the research results by Arif et al. (2019a) in Makassar, South Sulawesi, showed that pine wood bait planted for three months had not been attacked by subterranean termites, and termite attacks only occurred in the sixth month of observation. Therefore, it is necessary to research wood bait alternatives to pine based on species of community timber and find the optimal length of time for baiting.
2. MATERIALS and METHODS
The research was conducted from November 2020 to July 2021 at the Arboretum of the Faculty of Forestry and Environment, Bogor Agricultural University, Dramaga, Bogor Regency, West Java Province. The average air temperature in Bogor District is 21.1°C–33.7°C, the average humidity is 78%–89%, and the annual rainfall is 2,420 mm/year, with rainy days of 157 days/year (Central Bureau of Statistics, 2021). The termites were collected and identified at the Laboratory of Wood Quality Improvement Technology, Department of Forest Products, Faculty of Forestry and Environment, IPB University, Bogor, West Java, Indonesia. In addition, the soil of the research location was analyzed at the Indonesian Center for Biodiversity and Biotechnology (ICBB), Bogor.
Wood baits were installed randomly at the test site at the Arboretum of the Faculty of Forestry and Environment, IPB University, Dramaga Campus, Bogor Regency, West Java Province (Fig. 1). The plot size is a rectangle measuring 12 m (length) and 7 m (width). The test site coordinates are 106° 43’ 46.40”E, 6° 33’ 27.22”S; 106° 43’ 46.17”E, 6° 33’ 27.32”S; 106° 43’ 46.11”E, 6° 33’ 27.80”S, and 106° 43’ 46.43”E, 6° 33’ 27.74”S. The wood baits were buried until 25 cm of the total length of the wood was vertically immersed under the ground and the upper half of above ground the wood bait are and marked (Arinana et al., 2020). The distance between the wood bait in the longitudinal direction (row) was 30 cm, while the distance between the wood bait in the wide direction (column) was 60 cm.
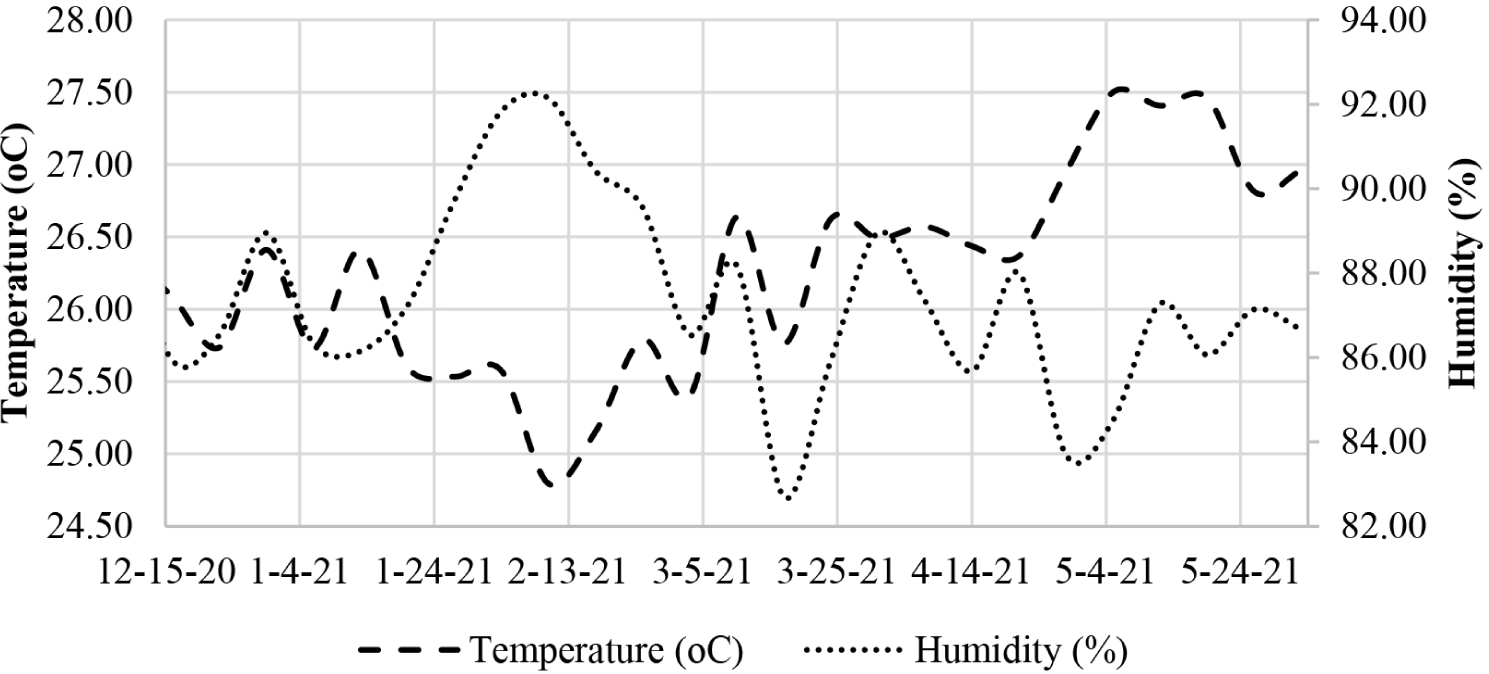
Temperature and humidity measurements were carried out using a Digital Temperature Humidity Data Recording Logger Meter type DWL-20 installed at one point around the test site. Temperature and humidity data were automatically recorded on the device every hour for six months.
Soil characteristic analysis was carried out following Arinana et al. (2016) to determine the suitability of the soil in the research area as a habitat for subterranean termites. Soil samples were taken at five locations: Four in the cardinal directions (east, west, south, and north) and one in the field test location’s center. At least 1 kg of soil from 0–25 cm depth was collected from each soil sampling station and placed in transparent plastics. The soil pH, C-organic content, and soil texture were measured at the ICBB Laboratory (an ISO/IEC17025 certified laboratory) in Bogor.
This study used logs of seven wood species with a diameter of 20–40 cm. The wood species were A. mangium, F. moluccana, A. cadamba, M. eminii, S. mahagoni, H. brasiliensis, and P. merkusii. All wood materials were collected from Ciherang Village, Dramaga, Bogor, West Java. This study used the wood part from the sapwood. The logs were then sawed into boards and wood samples (2 cm × 2 cm × 46 cm) with ten repetitions for each species following the American Society for Testing and Materials (ASTM) D 1758-06. The total of wood baits was 420 samples. The surface of the wood baits was sanded using sandpaper with grit number 100 to remove residual fibers from the cutting process, and the upper side was painted with a different color according to the length of baiting time. The wood bait was then kept in airflow (by a fan) until the moisture content was below 20% and weighed to acquire the initial weight before being installed in the research location.
The termite soldier caste specimens found on each wood bait were put separately into a collection bottle containing 70% alcohol. The termite specimens were identified, referring to Tho (1992) and Ahmad (1958). Morphological observations of termite specimens were carried out with the help of a digital microscope endoscope camera magnifier 800× 8 LED at a magnification of 10×.
Weight loss (WL) is the ratio between the decreasing weight of a sample (the initial weight (W1) minus the weight after baiting (W2) and the sample’s initial weight, expressed in percent (%). The calculation formula is as follows:
The best baiting time was determined by considering the wood baits’ weight loss percentage. Based on the JIS K 1571 (2010) standard, the test value limit is successful if the wood weight loss value is more than 15%.
The frequency of termite attack is the ratio between the amount of wood bait attacked by subterranean termites and the total amount of wood bait planted in the test area expressed in percent. According to Cookson and Trajstman (2002), the frequency of subterranean termite attacks is classified into six classes. The average termite attack frequency obtained in the test is compared with the data in Table 1.
| Frequency (%) | Category |
|---|---|
| 0 | No attack |
| 1–10 | Very low |
| 11–20 | Low |
| 21–30 | Moderate |
| 31–40 | High |
| > 40 | Very high |
The damage percentage was categorized into seven classes, from no attack to 75% cross-sectional damage (ASTM D 1758-06). Wood damage percentage (WDP) is the ratio between the length of the cross-sectional damage on the attacked wood sample (L) and the wood sample’s initial width (Wt), expressed in percent (%). The percentage of the cross-sectional damage determines the wood sample grading system on the wood sample due to the subterranean termite attacks (according to ASTM D 1758-06). See Table 2 for the wood sample grading system.
The data analysis used was a completely randomized design with one factor: The species of wood treatment or the length of time it was fed. Data were analyzed using analysis of variance (ANOVA) at the 5% test level using SPSS 25.0 software. If the ANOVA showed significant results, the analysis continued with Duncan’s test (Tallarida and Murray, 1987).
3. RESULTS and DISCUSSION
Scientists have been researching the interaction of termites with soil for many years. One of the most noticeable is the effect of soil on termite dispersal, although temperature and moisture play a significant influence (Cookson and Trajstman, 2002). Soil has an important impact on subterranean termites as a place to live and protects termites from extreme temperatures and humidity (Nandika et al., 2015). Termites selectively use soil particles, choosing finer particles and building structures that fit their ecological, physiological, and behavioral needs (Khan and Ahmad, 2018).
The soil texture is essential in building termite tunnels above or under the ground to connect main nests with food sources. Termites generally prefer habitats with clay soil texture and fine sandy intolerable (Lee and Wood, 1971). The results showed that the soil texture at the test site was clay with sand (10.3%), dust (13.8%), and clay (77.8%) and indicating that the study site is a suitable habitat. The low percentage of sand is preferred due to these types of soils having relatively low organic matter content. The values obtained are in close agreement with Kaschuk et al. (2006), the clay content in termite mounds is higher than in the surrounding soil. Therefore, clay has an important role in the formation of termite nests. The complete soil characteristics (pH, C-organic, sand, silt, clay, and sand/clay ratio) at the test site are presented in Table 3.
The results showed that the soil pH at the test site ranged from 4.54 to 4.78. According to research by Kaschuk et al. (2006), termites can survive in an acidic soil environment. Furthermore, based on the review of Li et al. (2017), most termites were nested in acidic and weakly alkaline soil with the highest frequency at 4.5 to 5.5. Soil acidity was determined by the organic matter content of the litter and the type of vegetation in the ecosystem. The main effect of termite activity is the ability to improve soil quality (local microsite). Therefore, termites are potentially an essential source of heterogeneity in soil systems in tropical forests (Arif et al., 2019a).
C-organic content is one of the determinants of soil fertility which functions to increase nutrients and as a nutrient buffer (Arif et al., 2019b). The content of C-organic in the soil is also beneficial for microorganisms in the soil, namely as a food source to meet their needs. The results showed that the C-organic content in the test site ranged from 1.91%–2.66%. The C-organic content in this research was lower than the results of Kaschuk et al. (2006).
According to the Indonesian Soil Research Institute (2009), the low-medium group classified the C-organic content. The C-organic content at the test site was higher than in the study site of Arinana et al. (2020), ranging from 0.69%–1.78%, with the test site in the form of settlements. According to Mahaney et al. (1999), termites play a role in increasing organic matter in the soil and is also supported by Jiménez and Decaёns (2006), stating that increased organic matter was obtained from the feces and excretion of termite saliva, which is the main component in building nests. The increase in organic matter in the soil will increase microbiological activity and population, especially in organisms related to the decomposition and mineralization of organic matter (Arif et al., 2019b).
Air temperature and humidity are the main factors that affect termites’ activity, development, and behavior (Ferreira et al., 2019). The results showed that the average air temperature at the study site was 25.15 ± 1.46°C, while the maximum temperature was 27.08°C and the minimum temperature was 24.62°C. Meanwhile, the average humidity reached 89.33 ± 4.44%, with a minimum humidity of 82.77% and a maximum humidity of 92.24% (Fig. 2). Based on Nandika et al. (2015), the temperature at the study site is the optimum temperature for termite activities. It is a suitable habitat for termite species because the temperature at that location was in the range of 25°C–28°C. According to Nandika et al. (2015), air humidity at the test site was included in the optimum humidity because the humidity is in the range of 75%–90%. Moisture is one of the factors needed by termites because termites need high humidity to prevent dehydration in termite nests.
The results showed that the termite specimens attacking the wood bait after exposure at the test site for one, two, three, four, five, and six months were mainly four species, namely Schedorhinotermes sp., Macrotermes sp., Microtermes sp., and Capritermes sp. (Table 4, Fig. 3 and Fig. 4). These subterranean termites are members of the Termitidae family, except Schedorhinotheres sp., a member of the family Rhinotermitidae. In the research of Hadi et al. (2018) at the same research site, where this research was conducted, the type of termite found was the Coptotermes curvignathus.
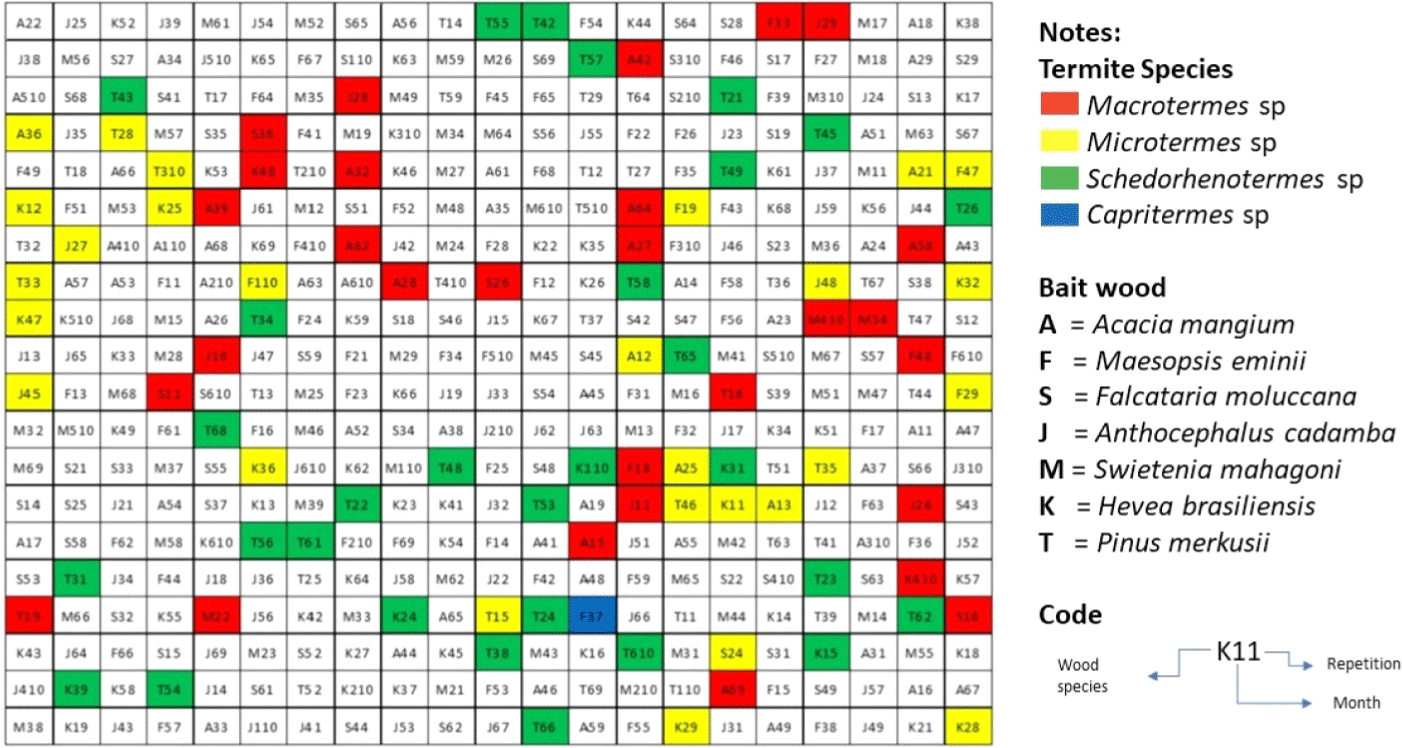

The data showed that termite species from the family Termitidae were more dominant than termite species from the family Rhinotermitidae. The dominance of Termitidae is due to their spectacular adaptation to sunlight radiation, comprising about 75% of termite fauna in the world (Krishna, 1970; Varley and Wilson, 1974). The Termitidae family belongs to the high-level termite group, which has three-fourths of the total termites and is mostly found in tropical areas (Nandika et al., 2015). Nevertheless, based on Mubin et al. (2016), the abundance of termite species Schedorhinotermes sp. was relatively numerous compared to other species of the family Rhinotermitidae in the plot site area. The presence of Schedorhinotermes sp. is more commonly found because these species have a wide range of foraging (Fig. 3).
In the first month of baiting, all subterranean termite species, except Capritermes sp. found to attack all species of wood bait, namely A. mangium (acacia), F. moluccana (sengon), A. cadamba (jabon), M. eminii (manii), H. brasiliensis (rubberwood), and P. merkusii (pine), except S. mahagoni (mahogany). Subterranean termites actively attacked wood bait from the first month to the sixth month on acacia and pine wood bait. Rubberwood and jabon wood bait were not attacked by termites in the 6th month. However, in the 5th and 6th months, they experienced significant damage; more than 80% of their weight was lost due to attacks. Based on species diversity, rubberwood and pine have a higher diversity of termites that attack the baits. Three species of termites attacked the species: Macrotermes sp., Microtermes sp., and Schedorhinotermes sp.. This result indicates that various species of termites prefer those species of wood.
In contrast, mahogany was only attacked by one species of termite and rarely attacked. Kalleshwaraswamy et al. (2022) said that different wood species have varying degrees of resistance to insect damage, like termite attacks. Based on termite species, Schedorhinotermes sp. attacked 30 wood baits but only rubberwood and pinewood species. Meanwhile, Macrotermes sp. found had actively attacked 29 wood baits and Microtermes sp. on 28 wood baits in the sixth month. These types of termites were also found to attack other wood species other than those tested in this study, such as the discovery of Macrotermes gilvus and Microtermes insperatus that attack teakwood plantations (Fajar et al., 2021), Schedorhinotermes javanicus that attack Acacia crassicarpa plantation (Haneda et al., 2017), and M. gilvus, Microtermes sp., and Schedorhinotermes medioobscurus that attack Eucalyptus pellita plantation (Prastyaningsih et al., 2020).
The results showed that 391 baits were attacked for six months of baiting time. Therefore, the frequency of termite attacks on wood bait at the plot site was 93.1%. During the observation, no active termites were found in some of the attacked wood bait. Subterranean termites had attacked all wood baits since the first month of baiting. The average termite attacks on all wood species ranged from 71.4%–100% (Fig. 5). After one month of baiting, acacia, manii, and rubberwood even had a termite attack frequency of 100%.
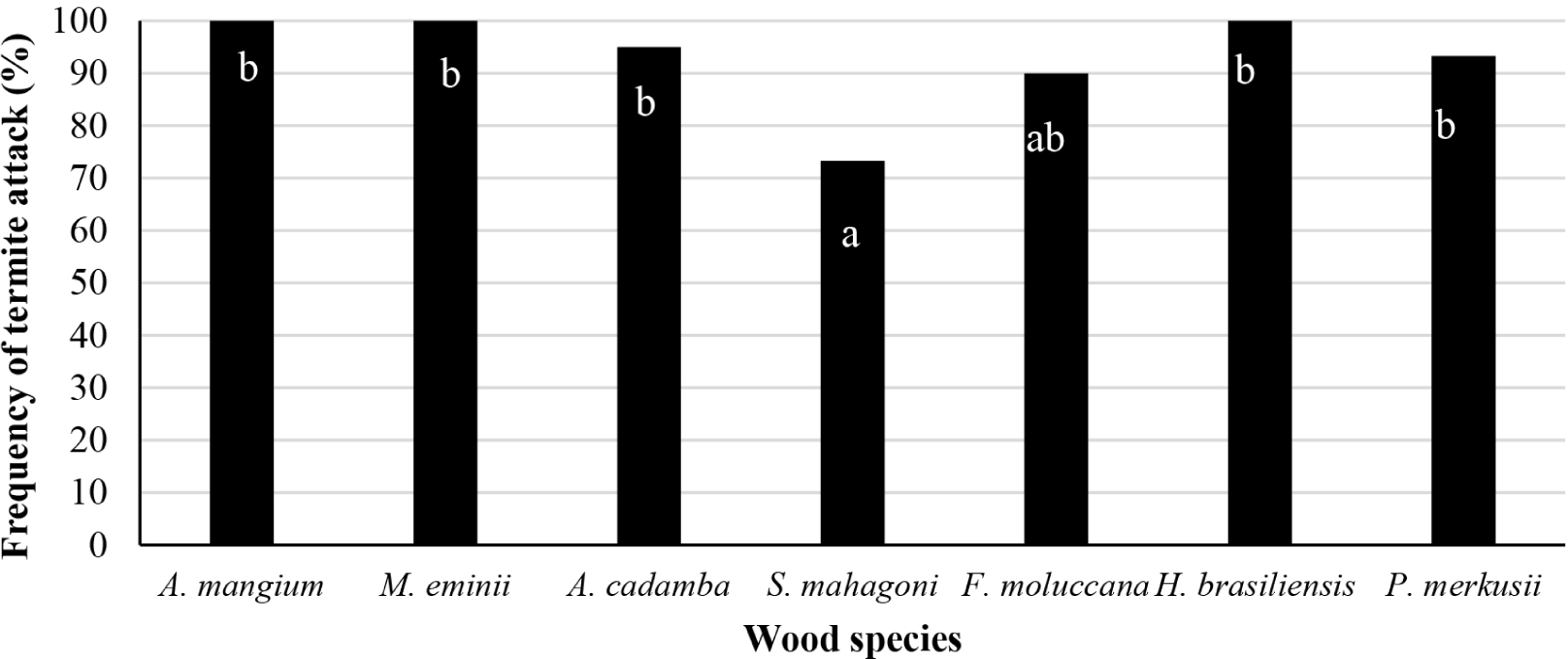
Meanwhile, subterranean termite attacks were 100% on jabon, sengon, and pine in the second month of baiting, while mahogany occurred only in the fifth month. The variation of baiting time was influenced by the level of durability of the wood being tested. The low level of wood durability makes the wood more susceptible to wood-destroying organisms, including termites. The results showed that the baiting time for two months is the optimal time that can be used as a baiting time in plot tests with the same characteristics.
The results showed that the weight loss of wood bait was observed in the first month of baiting (Fig. 6 and Fig. 7). Species and wood components caused the difference in weight loss in the wood bait. The significant weight loss in rubberwood was influenced by the nature of the wood, which is easily overgrown by fungi if the moisture content is more than 20%. Termites strongly favor moist and moldy wood conditions, making it easier for termites to attack the wood. This fact is in line with Viana-Junior et al. (2018), who stated that several termites prefer decayed wood. The fungus produces compounds that can degrade wood and attract termites to come and eat the bait. The low weight loss of mahogany was expected due to extractive compounds such as sesquiterpene and monoterpene produced from secondary metabolites that are toxic to termites (Deviriani et al., 2019). Chemical components of wood, especially extractive substances, are known to be factors that affect the natural resistance of wood, including attacks from subterranean termites (Aflah et al., 2021).
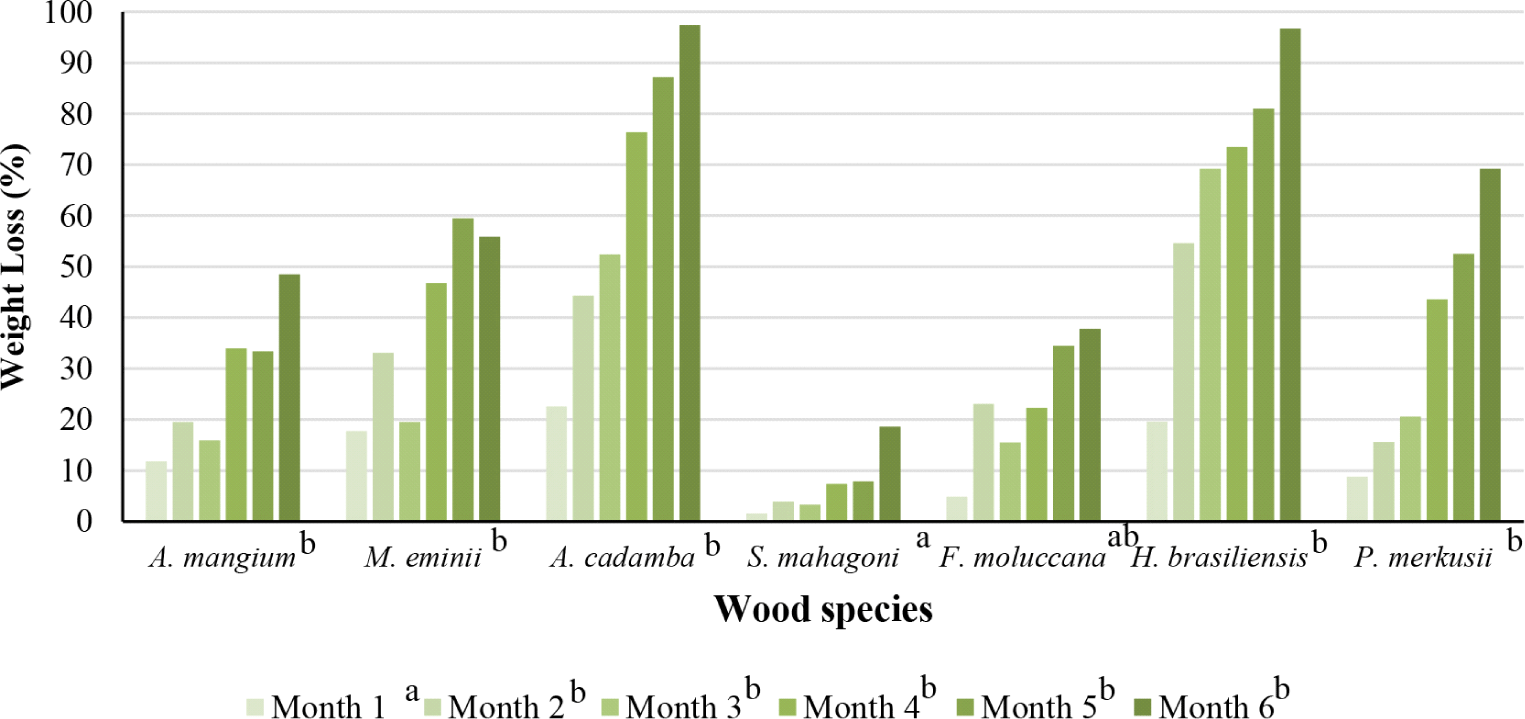

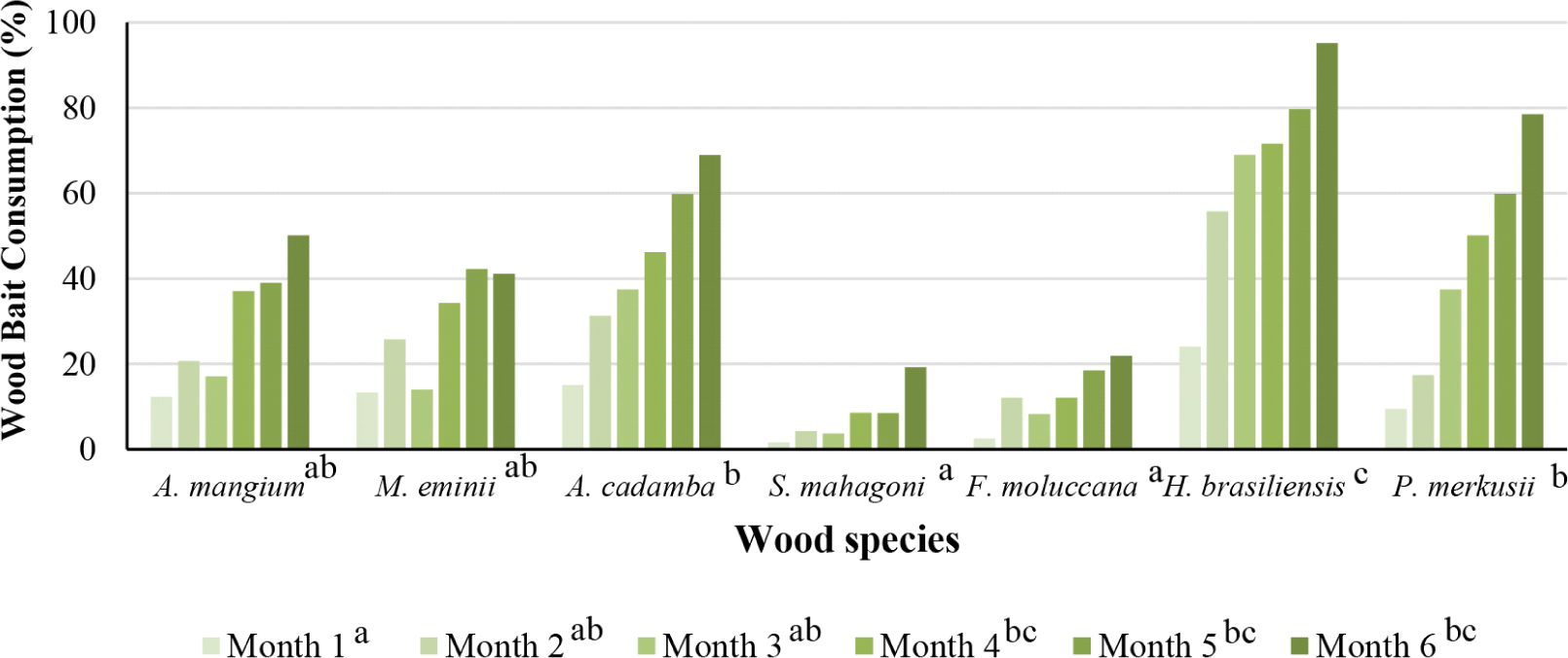
The consumption of wood bait by subterranean termites in the plot site has occurred one month since the wood bait was installed. In general, wood bait consumption increased with longer baiting time; even rubberwood consumption after six months of baiting was 95.2 g (average initial weight before baiting was 98.5 g), and at the end of the baiting, only 3.3 g. Rubberwood was the most consumed wood by termites, followed by pine and jabon. This finding showed that termites prefer rubberwood over pine (Fig. 8). The level of wood bait consumption by termites was influenced by several factors, namely environmental factors and colony size. Termite species that attack wood bait also affect the level of wood bait consumption (Subekti, 2012). The obtained data indicate that pine at the time of baiting has a relatively large average wood consumption. This finding was supported by Subekti (2012), that pine wood is the most preferred species of wood by subterranean termites M. gilvus. Rubberwood is also very susceptible to termite attacks. According to the results of this study, that rubberwood has a large weight loss (Grace et al., 1998).
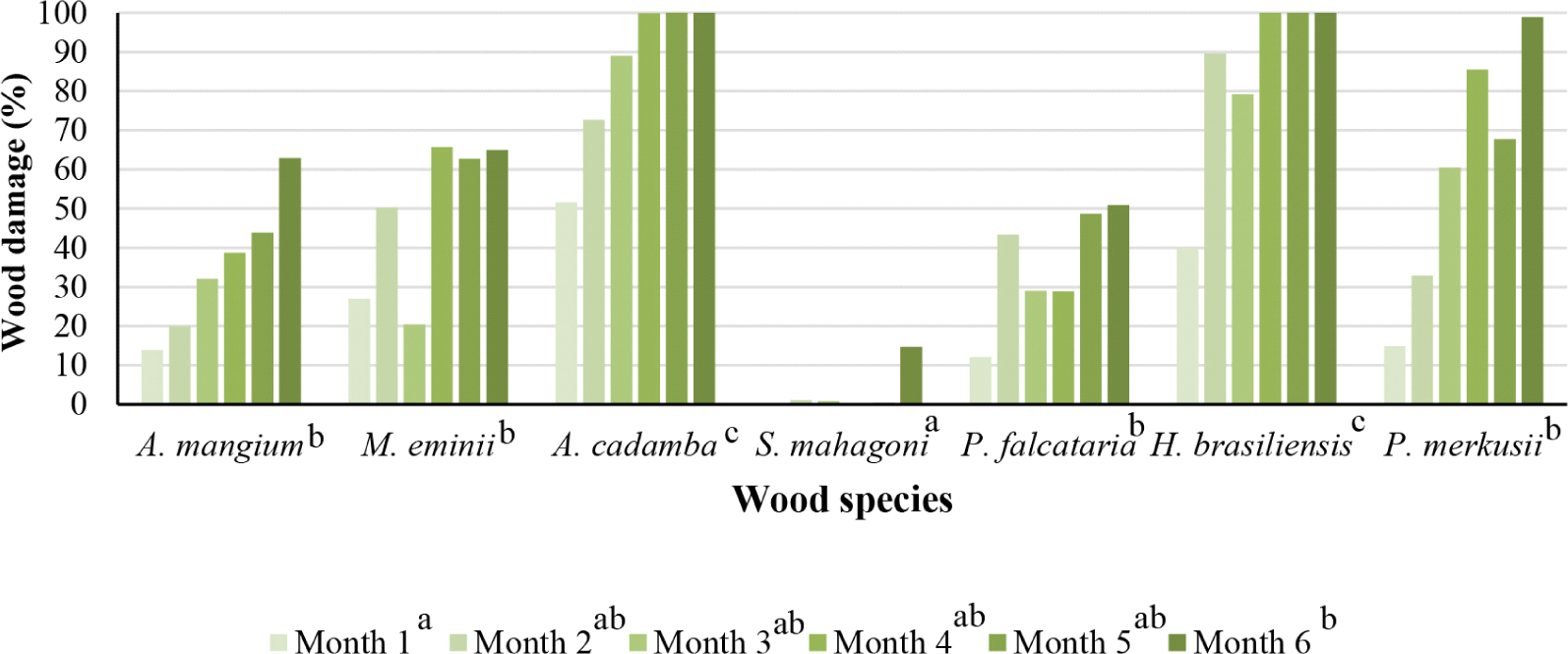
The percentage of damage in Fig. 9 showed that jabon wood and rubberwood were most favored by termites during the baiting, followed by pine, manii, acacia, and sengon wood. Mahogany had a relatively low percentage of damage in all baiting time ranges compared to the other six wood species. Rubberwood has the highest percentage of damage which means very low durability and according to SNI 7207:2014 (BSN, 2014), rubberwood was classified into durable class V. Meanwhile, Jabon was categorized as class IV due to its low resistance to subterranean termites (Arsyad et al., 2019). Acacia was classified as resistance class III and for sengon wood was class V (Hadi et al., 2020). The class also indicates that the wood species was favored by termites and caused greater damage. Based on the percentage of damage, jabon, rubberwood, manii, acacia, pine, and sengon were suitable for use as wood bait because these species tend to be favored by termites. Mahogany wood is not recommended for wood bait because, in all the baiting time, the wood had a relatively small percentage of damage, indicating that the wood was less favored.
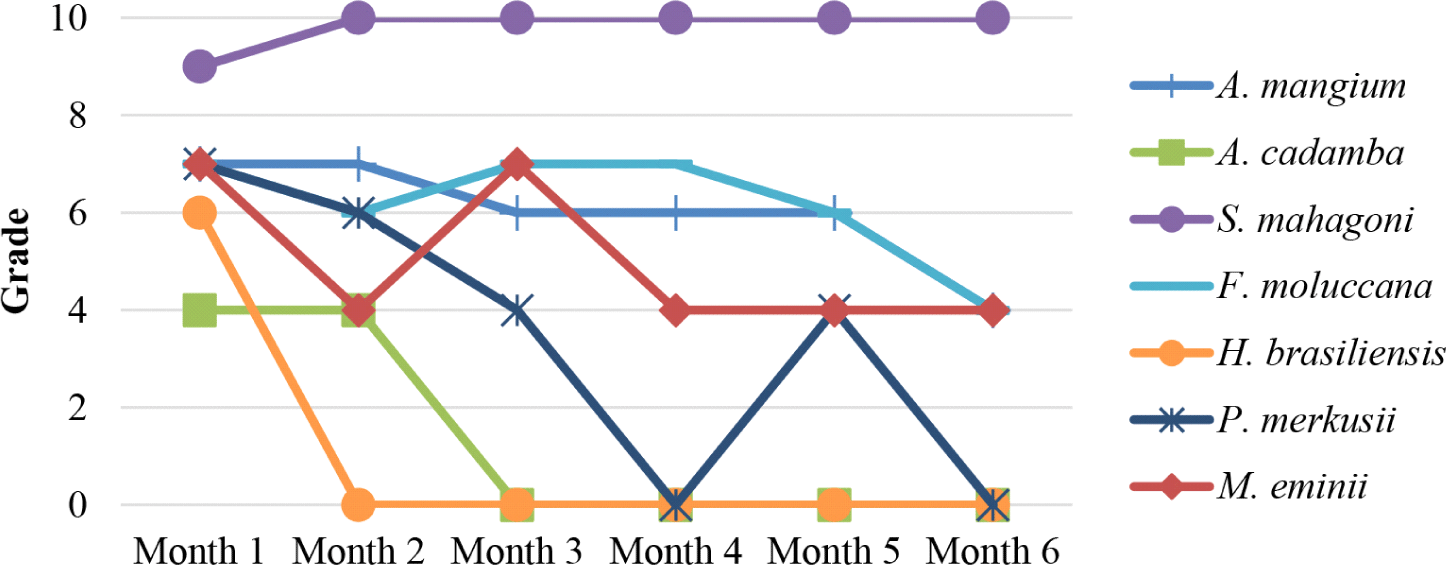
The results showed that the damage values of rubberwood, pine, and jabon wood bait were 0 (damaged) in almost all baiting time. The result showed that the percentage of damage to the wood was very high. Based on Fig. 10, rubberwood was drastically damaged. This finding showed that termites have a very high preference for rubberwood. The damage value showed that the damage value of the tested wood bait decreased relatively with increasing baiting time.
4. CONCLUSIONS
The weight loss value revealed the termite preference level in several observed wood species. The wood species providing favorable conditions to termites, termed as highly favored woods, have shown higher weight loss percentages and vice versa. The greater the weight loss percentage the more the wood was preferred by termites. The results have shown that termites have preferred rubberwood over other species. Rubberwood, jabon, and manii reached the minimum limit in a month. However, acacia, sengon, and pine have optimal baiting time in the second month.
Based on the results, it can be concluded that the order of the preferred wood species for the subterranean termites among community timber tested was rubberwood, jabon, manii, pine, acacia, sengon, and mahogany. Based on weight loss and frequency of attack, rubberwood, jabon, and manii have the potential to be an alternative to pine as a wood bait due to their susceptibility to termites and the shorter baiting time, usually up to one month.