
1. INTRODUCTION
Termite infestations in urban environments cause significant economic and structural damage worldwide. In temperate regions, infestations are usually dominated by a single genus, whereas in tropical regions, multigenera termites, consisting of lower and higher genera, infested buildings and other structures. The lower termites involved in urban infestations include Coptotermes, Heterotermes, Mastotermes, Prorhinotermes, and Schedorhinotermes (Lee et al., 2007; Manoppo et al., 2024). Furthermore, key higher termite genera that commonly infest tropical urban settings are Macrotermes, Odontotermes, Microtermes, Hypotermes, Globitermes, and Nasutitermes (Arinana et al., 2022; Lee et al., 2007). Conventional pest control strategies, often reliant on broad-spectrum liquid insecticides, have historically dominated termite management but present environmental concerns and practical limitations. This has led to a growing interest in alternative, more targeted approaches, such as baiting systems, which leverage the termites’ social foraging behaviors for colony elimination, rather than merely localized suppression (Chouvenc, 2025).
Commercial baits are effective at eliminating lower termites (family Heterotermitidae), especially the genera Reticulitermes and Coptotermes (Evans and Iqbal, 2014). However, in the tropics, reinfestation by other termite families usually occurs after successful elimination. The reinfestation is predominantly by Macrotermes, Microtermes, Globitermes, and Schedorhinotermes, which account for ~15% of total infestations (Lee et al., 2007). Furthermore, Chiu and Li (2024) found that higher termites do not respond well to baits made from a pure cellulose-based matrix because they have broad dietary preferences. This affects the efficacy of the bait system in controlling the termite population, resulting in a long interception time, which increases the control time and costs.
The success of baiting systems depends on several factors, such as the active ingredients, palatability, and attractiveness of the bait matrix. An unsuitable bait matrix is more likely to be abandoned if termites encounter more suitable food sources or if disturbances occur at the bait stations (Swoboda, 2004). In general, termite food selection based on the production of a bait matrix is influenced by the degree of wood decay, nutritional content, secondary metabolites, and fatty acids that affect termite feeding and survival (Arinana et al., 2024; Judd, 2018). Using a wood-bait matrix preferred by multigenera subterranean termites, such as ex-tapped rubberwood, is a potentially effective way to transform agricultural waste into a value-added solution for termite control in tropical regions.
Although subterranean termites tend to feed on wood previously decayed by specific fungi, they can also attack fresh wood. In some cases, fresh wood stakes perform better than commercial bait (Pawson and Gold, 1996), which made the wood-based matrix a promising matrix. Another strong argument for wood-based matrix is their durability, which allows for a long feeding period for termites to ensure a colony-wide distribution of toxicants.
Subterranean termites generally exhibit a strong feeding preference for pines (Pinus spp.; Judd, 2018). Other types of wood that are preferred by termites are rubberwood (Hevea brasiliensis), jabon (Antocephalus cadamba) and African wood (Maesopsis eminii; Arinana et al., 2022). Rubberwood is attacked by a wider range of termites and showed a similar feeding preference to that of pine. Based on the level of infestation and weight loss, rubberwood was higher than that of the other two species (Arinana et al., 2022). Therefore, with a variety of termite species attacking rubberwood baits, the high frequency of attacks and weight loss rates suggest that rubberwood could serve as a versatile bait matrix for multigenera subterranean termites.
Despite this potential, the underlying factors that make rubberwood attractive to multigenera subterranean termites remain poorly understood. Detailed studies on termite preferences, including physico-chemical properties, such as wood hardness and extractive content, are needed to identify the attributes responsible for its suitability as bait. Therefore, the objectives of this study are to explore the potential of rubberwood as a bait matrix for multigenera termites in the tropics. Specifically, we hypothesized the following:
-
Termite preference for rubberwood is influenced by its physical (e.g., hardness) and chemical properties.
-
The extractive compounds in rubberwood act as attractants and phagostimulants, enhancing feeding and shortening bait interception times.
By addressing these hypotheses, this study aimed to fill a critical research gap in bait matrix development for tropical termite control by integrating physico-chemical wood traits, gas chromatography-mass spectrometry (GC-MS)-based semiochemical profiling, and field interception performance across multigenera subterranean termites.
2. MATERIALS and METHODS
A preference test was conducted using three wood types: rubberwood (H. brasiliensis), jabon (A. cadamba), and acacia (Acacia mangium). The bait was made from sapwood cut into stakes with dimensions of 3 × 3 × 35 cm [Fig. 1(a)]. The stakes were oven-dried at 60°C until they reached a stable weight. Before installation in the field, the stakes were soaked in distilled water overnight to restore their moisture content.
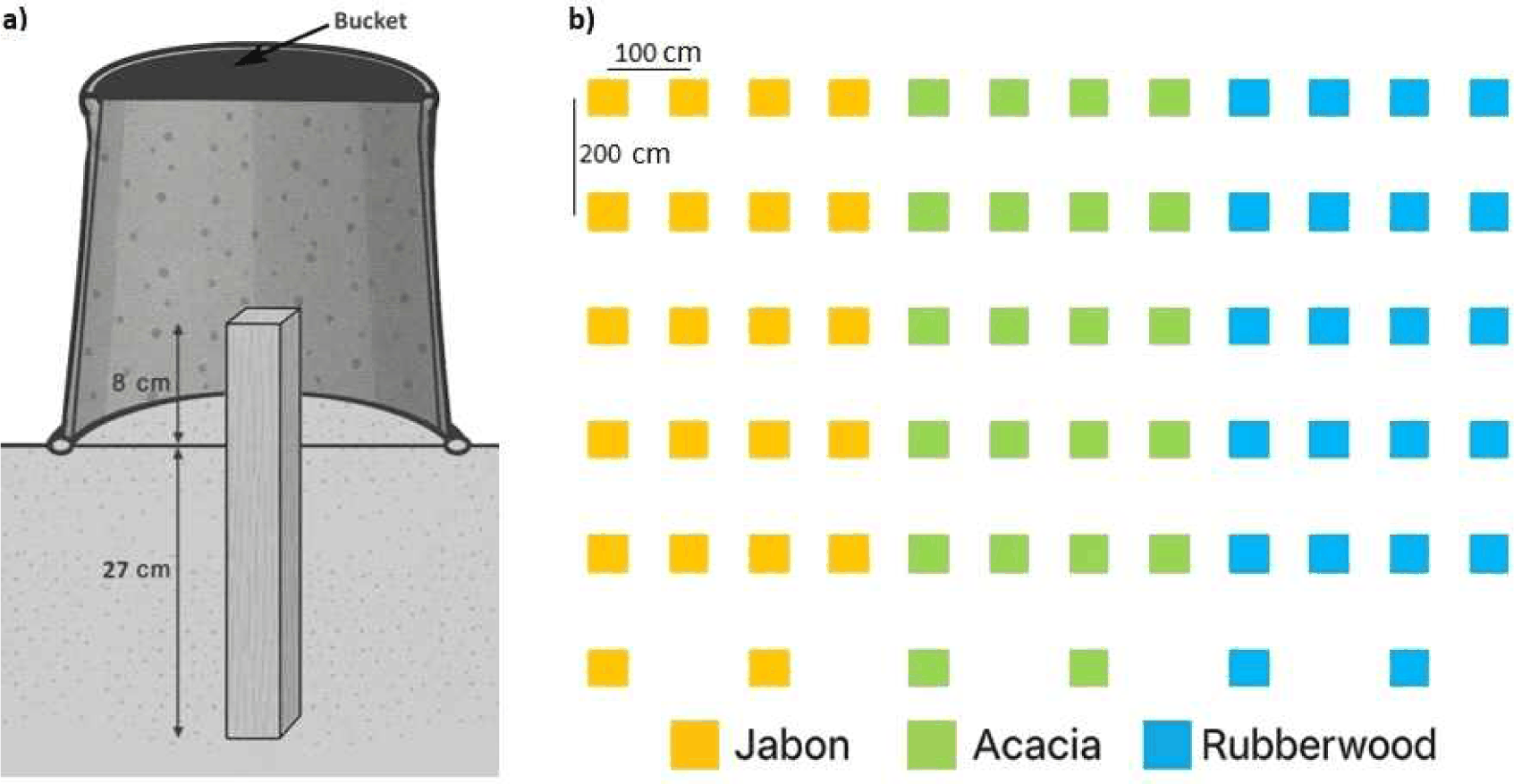
The preference test was performed in the Cikampek Special Purpose Forest Area (KHDTK Cikampek), Karawang, West Java, Indonesia, using a grave-yard test method. Stakes were installed vertically; 27 cm of the stakes were on the ground, and the remaining 8 cm remained above the soil surface [Fig. 1(a)]. There were 22 stakes for each wood species installed in 11 × 10 m plots in species-specific groups within a single continuous test area. Grouping by species was applied to avoid cross-attraction and ensure that feeding activity on each wood type reflected termite responses to that substrate rather than interactions among different woods.
The distance between the stakes was 1 m between rows and 2 m between columns [Fig. 1(b)], and then covered using a black bucket [Fig. 1(a)]. The code for each sample was assigned to the end of the stakes above the soil surface, as well as the bucket that covered the stakes. Monitoring of infested wood was carried out monthly until the end of the study (3 months). Termite-infested stakes. were observed from soil sheet building activity on the above-ground parts of the stakes. The soil moisture was monitored monthly using a soil tester.
All the stakes were collected at the end of the study. Termites found on the stakes were then preserved in a container containing 70% of alcohol for identification. The subterranean termite genus was identified based on the morphology of soldier castes using a stereomicroscope. Identification refers to the identification key of Termites of Peninsular Malaysia (Tho and Kirton, 1992). The stake wood was then oven-dried at 60°C until it reached a stable weight and cleaned. It was then weighed to determine the difference in weight, which represented the amount of wood consumed, using Equation (1):
where: WL = weight lost (g); W1 = dry weight of wood stakes before preference test (g); W2 = dry weight of wood stakes after preference test (g).
To gain clearer insight into how long it takes for rubberwood stakes to become infested, a bait station setup was employed after the preference test. The bait station allowed for more accurate monitoring of termite infestation on stakes by opening the bait cap and observing the presence of foragers or soil sheet building activity. Ten bait stations were installed, and rubberwood stakes with dimensions of 3 × 3 × 15 cm were inserted [Fig. 2(a)]. The inspection was performed every 10 days. If there was a sign of termite infestation in the form of a forager or soil sheet building activity, it was marked as a bait station infested by subterranean termites.
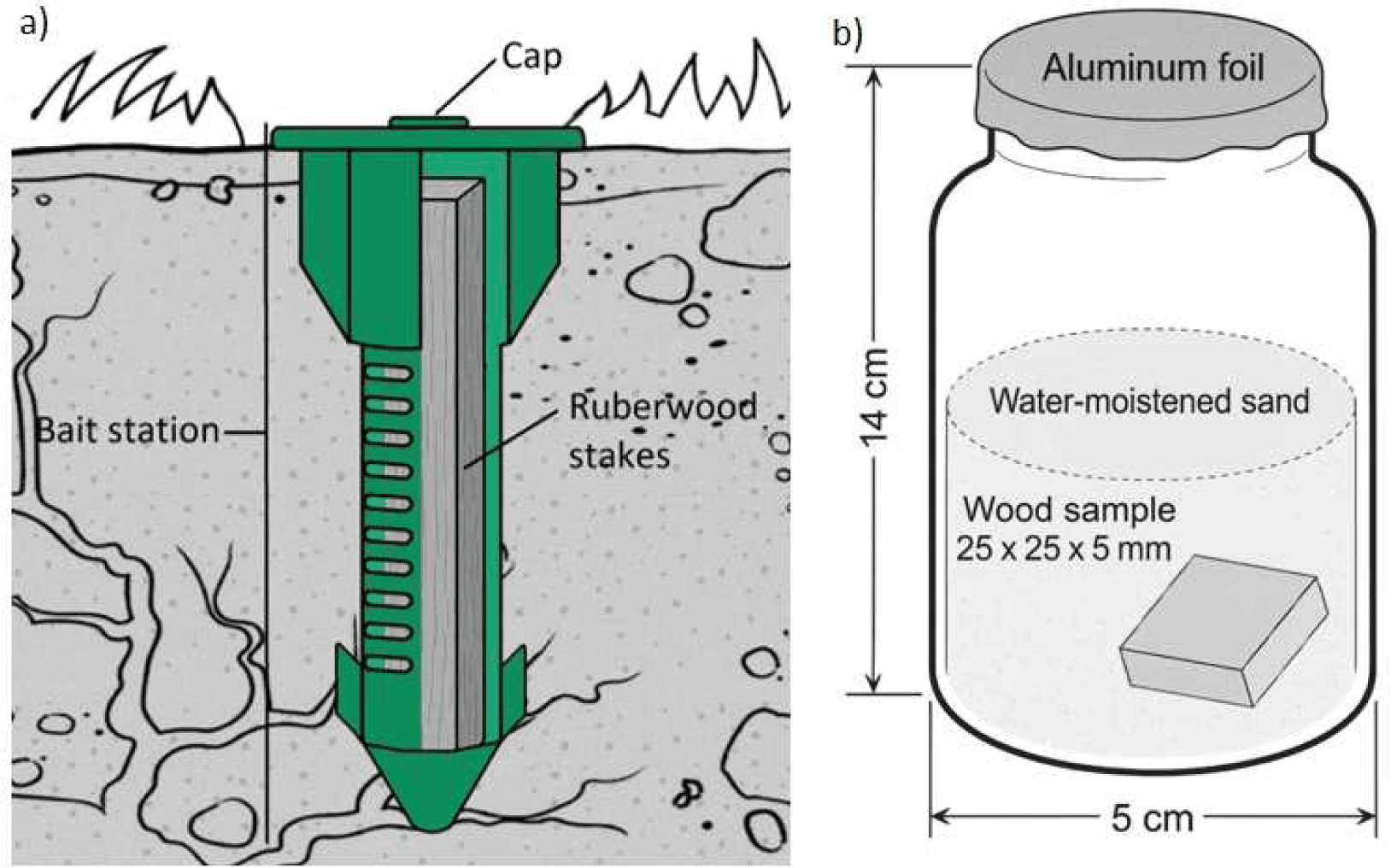
Further exploration of the potential of rubberwood as bait was carried out with the no-choice test, referring to the Indonesian National Standard (SNI) 01.7207-2006 method (SNI, 2006). Rubberwood samples treated with steam, boiling, and high-pressure steam were tested. Steam and boiling treatments were given to the stakes with durations of 1, 3 and 5 h at a temperature of ± 100°C. High-pressure steam treatment was carried out using an autoclave with the same duration as steam and boiling treatment with a pressure of 1 bar and a temperature of ± 110°C. The treated rubberwood stakes were then cut into 25 × 25 × 5 mm pieces, which were used as samples in the palatability test.
The palatability test was conducted using a 14 × 5 cm jar. In each jar, a wood sample (25 × 25 × 5 mm) was placed in an inclined position, with one side touching the bottom and the other side touching the jar wall [Fig. 2(b)]. It was then filled with 200 g of sterile sand, and 50 mL of distilled water was added. Termites from Coptotermes curvignathus were added to a jar with a composition of 200 workers and 20 soldiers for each sample (jar), covered with aluminum foil, and stored in a dark room for 4 weeks, with five replications for each treatment.
At the end of the test, wood samples were cleaned and oven-dried at 60°C until they reached a stable weight. The samples were weighed to determine the percentage weight loss, which represents the amount of wood consumed, using Equation (2).
where: WL = weight lost (%); W1 = dry weight of wood sample before test (mg); W2 = dry weight of wood sample after test (mg).
The physico-chemical of the rubberwood was performed on the control and treated samples. The wood hardness was tested with the Janka hardness test perpendicular to the fibers using a universal mechanical test instrument, referring to BS-373-1957 (BS, 1957). Hemicellulose and α-cellulose content measurements were based on the browning method (Browning, 1967). Insoluble lignin measurement was determined by the Klason method (Tappi T 222 om-88; TAPPI, 1994). The chemical content of the wood was determined using ethanol-benzene solvents. Volatile compounds in rubberwood were analyzed using GC-MS (Agilent Technologies, Waldbronn, Germany) with methanol as the solvent.
Descriptive statistics were used to show wood consumption and other parameters; all results in the main text are presented as mean ± SD. Statistical analysis was conducted using ANOVA, followed by Tukey’s honestly significant difference (HSD) post hoc tests, where appropriate, to determine the significance of wood consumption. Principal component analysis (PCA) was conducted to determine the physico-chemical factors of rubberwood that most influenced palatability. All statistical analyses were performed using Minitab version 21.4.3.
3. RESULTS and DISCUSSION
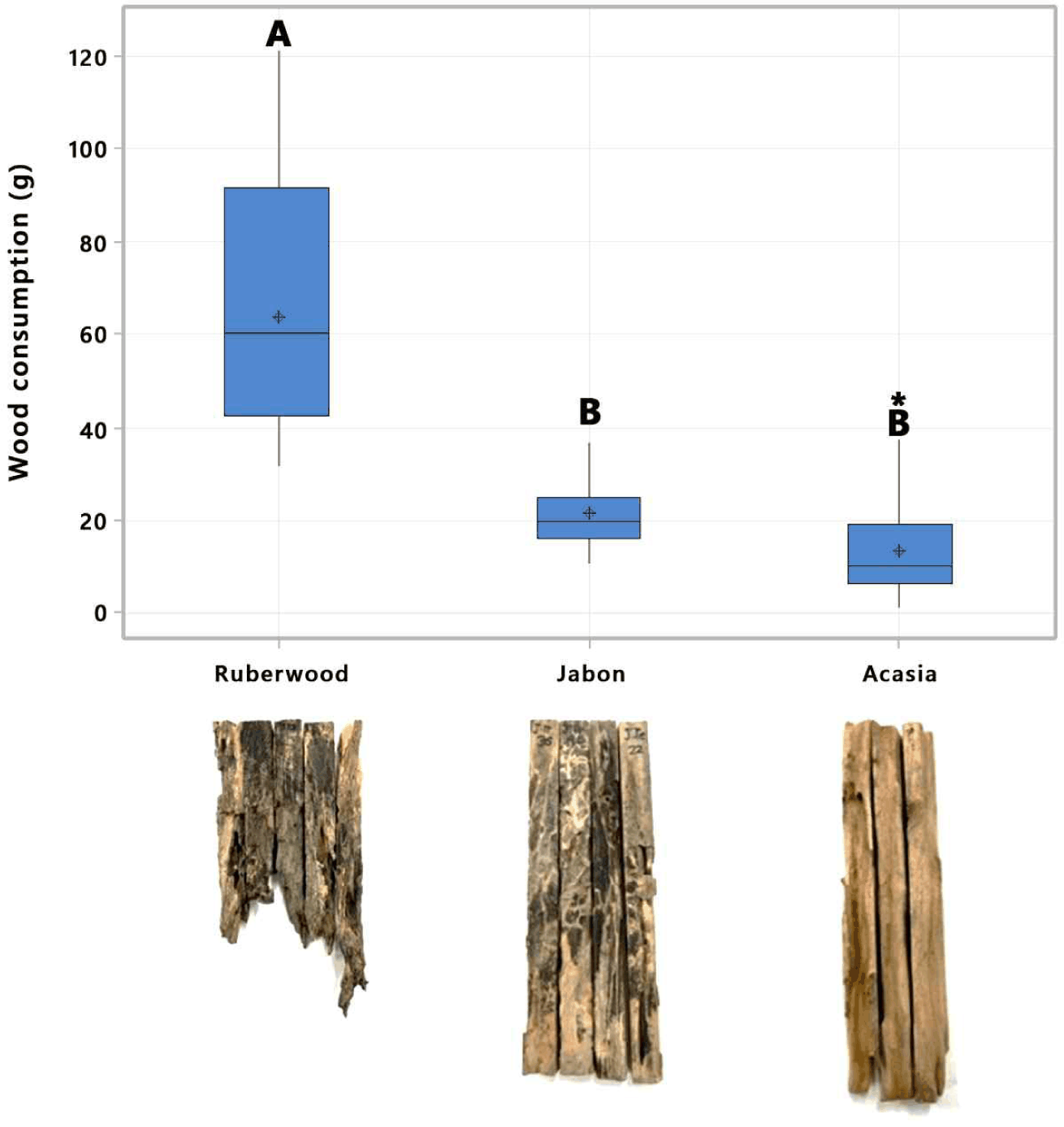
Preference tests revealed significant differences between rubberwood, jabon, and acacia stakes (F2,63 = 52.01, p = 0.00; n = 66). Rubberwood consumption was 67.2 ± 29.5 g (45.35 ± 21.7%), higher than jabon (21.7 ± 5.1 g) and acacia (15.2 ± 11.1 g; Fig. 3). The high consumption rate of rubberwood indicates that subterranean termites prefer bait made from rubberwood over that made from other wood species. Damage to rubberwood stakes due to subterranean termite investment is also extensive compared to jabon and acacia stakes (Fig. 3). This result is in line with the findings of Arinana et al. (2022), as rubberwood stakes tested in a residential environment had the highest weight loss (65.8%) compared to other commercial wood species.
The high consumption of rubberwood indicates that it is a quality food source; therefore, foragers that find the bait leave a pheromone trail to recruit other worker termites to consume the bait. As more workers consume bait, more of the bait matrix will be transferred to the colony members. This is an important part of termite control using bait systems, which exploit the foraging and food distribution behaviors of subterranean termites to deliver active ingredients throughout the colony (Su et al., 1995).
To achieve effective control with the bait system, subterranean termite foragers must find and recruit other foragers to the bait (interception time) and consume a sufficient amount of active ingredients that are transported back to the nest and distributed to other colony members (bait toxicant acquisition time) and the time required for the toxicant to kill termites (lethal time; Su, 2019). With a larger number of foragers consuming bait, the time required for termites to acquire a lethal dose of bait is expected to decrease.
Subterranean termite species that attack rubberwood are also high, which is 75% of the genera in KHDTK Cikampek. The genera that attack rubberwood consist of Coptotermes, which is a lower termite; Microtermes and Nasutitermes, which are higher termites. The high preference for rubberwood in the tropics is due to its high sugar and starch content, which makes it highly palatable to these insects (Chotikhun et al., 2024). Several studies in Indonesia and Malaysia demonstrate various termite species, including Microtermes sp., Macrotermes sp., Shedorhinotermes sp., and Capritermes sp. (Arinana et al., 2022), as well as Coptotermes gestroi, C. curvignathus, Globitermes sulphureus, and Microcerotermes crassus (Ngee et al., 2004). This preference from lower and higher termite species and high bait consumption makes rubberwood an effective baiting matrix for multigenera subterranean termites in the tropics compared to other wood species, including pine, which has been commonly used as bait.
The duration of stakes infested by termite infestation was monitored monthly by monitoring the activity of the soil sheet built on the stakes [Fig. 4(a–c)]. In the first month, jabon was the most highly infested at 58%, followed by rubberwood at 45%, and acacia at 38%. In the second month, rubberwood baits were the most highly infested (68%), followed by jabon (63%), and acacia (58%), as well as in the third month. The higher initial attack rates observed on jabon during the first month were likely influenced by its spatial proximity to termite nests or established foraging paths within the field site, which can strongly determine early bait encounter rates. However, the subsequent shift in feeding activity, with rubberwood exhibiting higher attack intensity and greater consumption in the second and third months, indicates that factors beyond the initial encounter, particularly the physico-chemical properties of the wood, may play a stronger role in sustained or later-stage feeding rather than initial discovery (Ali et al., 2021).
Bait discovery duration tests with the bait station system showed that 40% of rubberwood stakes had been infested by termites within 10 days after installation, with one of the bait stations having > 50% damage to the stake [Fig. 4(d)]. The short duration of bait discovery may have been influenced by bait cues. Cues that help termites find food sources include thermal shadows and chemical compounds produced by decomposers (Judd, 2018). Rubberwood is easily colonized by fungi such as Paecilomyces maximus, P. formosus, P. lecythidis,Penicillium crustosum, and Aspergillus chevalieri (Nandika et al., 2023), especially in humid areas. Fungal activity produced carbon dioxide gas, ions, and volatile compounds as chemical signals (cues) that attract foragers to the bait.

Baiting was carried out at the beginning of the rainy season, which increased soil moisture from 12.5 ± 7.5% when the bait was installed to 15.0 ± 0.0% (first month), and 57.5 ± 5.0% (second month). Increasing soil moisture alters foraging behavior, as foragers are more likely to forage near the soil surface, which improves the probability of detecting cues from bait and shortening bait discovery time (Sutherland et al., 2022). According to Cornelius and Osbrink (2011), increasing soil moisture can increase wood consumption, triggering fungal growth on the wood that allows foragers to detect chemical cues from baits. This study shows that application time (early rainy season) and bait material (easily decomposed wood) strongly affect bait success.
Although early baiting systems relied on a wooden matrix, modern commercial alternatives have largely transitioned toward a synthetic or durable matrix (Su, 2025). This shift is driven primarily by the susceptibility of wood to soil degradation, which necessitates frequent inspections and replacements, thereby inflating maintenance costs. From a purely financial perspective, fast-decaying wood such as rubberwood is often viewed as less viable than synthetic alternatives. However, in tropical regions, this rapid decay is beneficial, as the breakdown process releases chemical signals that effectively guide foraging termites toward the bait station. This biological signaling offers a trade-off as a distinct advantage over commercial baits that remain inert and undetectable in the soil for years.
In many regions, the time termites takes to intercept a bait station is the primary delay in the control process (Jones, 2003). However, this study demonstrated that rubberwood can shorten this window. Given the relatively shorter interception time in the tropics, the high palatability of rubberwood can significantly reduce the overall duration of the control process.
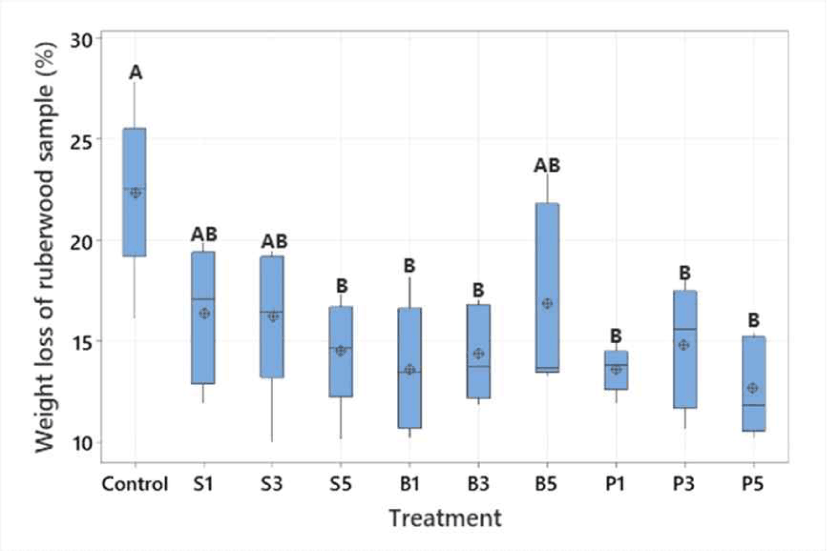
The highest palatability was recorded in the untreated sample (control), with percentage of weight loss is 22.36 ± 4.18%. Application of steam, boiling and high-pressure steam treatments (1, 3 and 5 h) showed significantly difference level of rubberwood consumption, ranging from 16.83 ± 4.65% to 12.67 ± 2.40%, among treatments (F9.40 = 3.63, p = 0.002; n = 50; Fig. 5). This shows that all treatments decreased termite feeding, suggesting that these treatments enhance wood resistance by reducing the palatability for termites. Similar results were also found for pine wood stakes, as heating treatments (steamed, oven, boiled, and high-pressure steam) significantly reduced termite-induced weight loss, indicating an increase in wood resistance (Arinana et al., 2020). This is because the heating treatment caused chemical changes in wood, particularly in hemicellulose, lignin, and the extractive (Park et al., 2018; Priadi et al., 2023). These results indicated that untreated rubberwood has the best palatability as bait for subterranean termites.
To determine the physico-chemical properties of rubberwood that correlate with palatability, PCA was conducted. The PCA extracted two major components explaining 75.0% of the total variance (Fig. 6). PC1 accounted for 46.6% and was mainly associated with hemicellulose, hardness, lignin, α-cellulose (negative), and ethanol–benzene extractives. PC2 contributed 28.4% of the variance and was dominated by lignin (positive), as well as hardness, and ethanol–benzene (negative loadings). In PC2, the consumption was negatively correlated with lignin and hemicellulose, and an increase in these compounds resulted in a decrease in consumption. The α-cellulose showed a less clear relationship to consumption, indicating that cellulose content alone is not a limiting factor in the consumption. The extractive substance content and wood hardness were closely related to rubberwood consumption. Overall, the PCA showed that lignin and hemicellulose contents were the strongest determinants of wood consumption, whereas extractives and hardness played secondary roles.
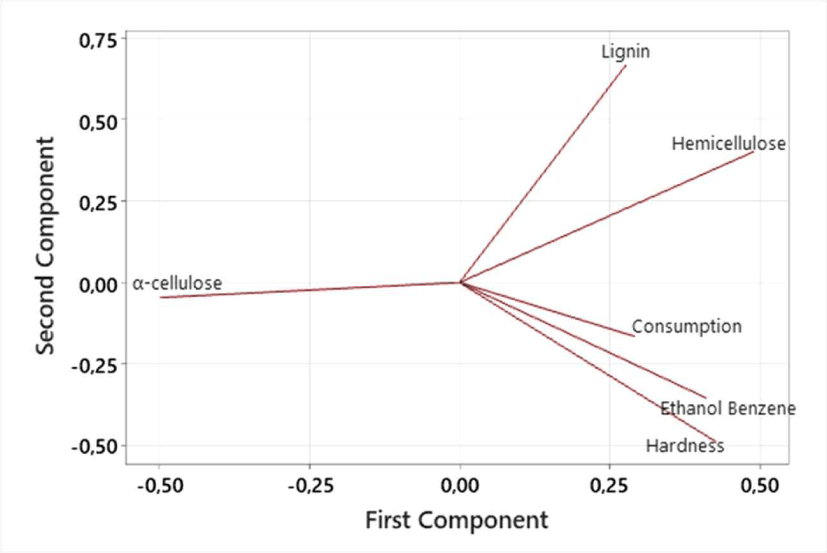
Hemicellulose and lignin were negatively correlated with consumption, suggesting that higher contents of these components reduce feeding activity. This is consistent with the structural and chemical roles of hemicellulose and lignin in enhancing cell-wall rigidity and reducing digestibility. Hemicellulose, although more readily degradable than cellulose, often interacts with lignin through cross-linking (Scheller and Ulvskov, 2010), forming a matrix that limits enzymatic access to cellulose, thus decreasing the palatability for termites.
However, there were differences in the abilities of the lower and higher termites to degrade lignin and hemicellulose. Li et al. (2023) showed that lower termites have a less ability to degrade lignin than higher termites. It is also the case with hemicellulose, where higher termites can digest hemicellulose as efficiently as cellulose (Xue et al., 2024). This is because the bacterial symbionts in higher termites are more diverse, producing endo-xylanase, acetyl xylan esterase, and mannanase enzymes that facilitate the comprehensive degradation of complex polysaccharides. Therefore, the observed negative correlation between lignin/hemicellulose and consumption is likely driven primarily by lower termite species (e.g., Coptotermes), which are generally less efficient at degrading the lignocellulosic complex. The successful consumption of rubberwood by both lower and higher termites, a key finding for multigenera baits, indicates that this matrix is broadly acceptable across functional groups. Although certain structural components of rubberwood may deter some lower termites, the presence of highly preferred extractives and the superior wood-processing ability of higher termites (e.g., Microtermes and Nasutitermes) appear to offset this effect, resulting in overall high consumption.
In contrast, α-cellulose levels showed no clear correlation with consumption. This indicates that cellulose content alone is not a limiting factor for termite feeding. Although termites possess cellulolytic symbionts capable of digesting crystalline cellulose, previous studies from Lee et al. (2020) reported that increasing the amount of cellulose substrate resulted in only minimal changes in the production of reducing sugars. This suggests that the enzymatic activity of cellulose is low because its high-molecular-weight polysaccharide structure is difficult to break down into simple sugars. Castillo et al. (2013) showed species-specific preferences for sugar supplements, with C. curvignathus preferring glucose-treated materials and C. gestroi favoring xylose treatments over pure cellulose baits, further supporting the importance of non-cellulosic nutrients in termite feeding behavior.
Wood, though rich in carbohydrates, is nitrogen-poor (0.56 ± 1.97% N; Maiti et al., 2019). For higher termites (e.g., fungus-growing termites), a bait matrix composed purely of cellulose may not be the most palatable diet (Chiu and Li, 2024). They also forage a variety of food sources like tree bark, dead branches, rotten wood, leaf litter, fruits, algae and animal dung, to provide nitrogen intake (Chiu et al., 2019). This indicated that the performance of rubberwood-based baits may be reduced in communities dominated by fungus-growing termites (Macrotermitinae), likely because of their distinct foraging ecologies.
The grouping of extractives and hardness in PC2 highlights their close relationship (Fig. 6). Although rubberwood is classified as a hardwood, Jung et al. (2025) reported that hardness alone does not guarantee resistance to termite activity because wood type is not a consistent indicator. They further emphasized that hardwood species vary widely in their vulnerability, revealing that their susceptibility to termite damage is far from uniform.
The rubberwood Janka hardness test values range from 3,189–4,688 N with an average of 4,029 ± 701 N. A study by Peralta et al. (2004) showed that the wood-consumption rates of forest subterranean termite species were not significantly correlated with wood hardness. This is also evident from a study conducted by Morales-Ramos and Rojas (2001), where wood preferred by termites had a wide range of hardness in the range of 2,870–8,200 N, whereas wood that induced feeding deterrence had a hardness in the range of 1,560–12,000 N. This information suggests that termites can consume wood with a fairly wide range of hardness, and it appears that the extractive substance content is an important component that correlates with rubberwood consumption.
Together, these results suggest that the palatability of rubberwood by termites is not solely determined by cellulose content but is also strongly influenced by the protective role of lignin–hemicellulose complexes, wood mechanical properties, and extractive compounds. Lignin acts as the chief physical and chemical barrier preventing enzymatic access to polysaccharides. Consumption was positively correlated with hardness and ethanol–benzene extractives. This suggests that termites do not strictly avoid high-density wood and indicates the presence of chemical signals (cues/stimulants) that are stronger than the physical resistance of wood (Etcheverry et al., 2025). This indicated a compensatory feeding strategy.
GC-MS analysis of the methanolic rubberwood extracts revealed six volatile constituents belonging to the lactone, phenolic, and fatty acid groups (Table 1). These metabolites have been widely reported in insect and termite chemical ecology and are known to function as attractants or phagostimulants. All compounds were consistently detected across the GC-MS analyses, confirming qualitative reproducibility; however, their relative abundances (% area) may vary among individual rubberwood samples owing to natural intraspecific chemical variability. Collectively, the presence of these compounds in combination with the physical properties of rubberwood may contribute to sustained feeding.
| Chemical compound | Group of compound | RT (min) | Area (%) | Bioactivity | Species/organism |
|---|---|---|---|---|---|
| 2H-Pyran-2-one, 5,6-dihydro-6-pentyl | Lactone | 22.102 | 0.22 | Attractant |
Thrips obscuratus (New Zealand flower thrips; El-Sayed et al., 2014) Cockroaches(Periplaneta spp.; Szőri et al., 2008) |
| trans-Isoeugenol | Fenol | 27.025 | 0.12 | Attractant |
Diabrotica barberi(northern corn rootworm; Whitworth et al., 2002) Zeugodacus diversus(fruit fly; Royer et al., 2018) Coptotermes curvignathus(Achmad et al., 2021) |
| Arachidonic acid (5,8,11,14-Eicosatetraenoic acid, methyl ester, (all-Z)-) | Fatty acid | 38.431 | 0.16 | Eicosanoid precursor | Insects(general; Vatanparast et al., 2018) |
| Developmental regulation | Insects(general; Vatanparast et al., 2018) | ||||
| Essential nutrient | Insects & termites(Vatanparast et al., 2018) | ||||
| Methyl palmitate (hexadecanoic acid, methyl ester) | Fatty acid | 38.634 | 0.32 | Attractant | Varroa jacobsoni(parasitic mite; Le Conte et al., 1989) |
| Kairomone(probing cue) | Peregrinator biannulipes(Tebayashi et al., 2003) | ||||
| Antibacterial | Termite (Arinana et al., 2025) | ||||
| Acaricidal | Tetranychus spp.(spider mites; Wang et al., 2009, 2010) | ||||
| Growth inhibition | Crickets(McFarlane and Henneberry, 1965) | ||||
| Repellent(short-chain fatty acids) | Stored product pests(Cohen et al., 1974) | ||||
| Methyl oleate (9-Octadecenoic acid (Z)-, methyl ester) | Fatty acid | 40.861 | 0.22 | Communication cue | Macrotermes gilvus(Kuswanto et al., 2015) |
| Oleic acid | Fatty acid | 41.225 | 0.12 | Burial cue(corpse burial) | Coptotermes formosanus(Li et al., 2024) |
| Increased building behavior | Reticulitermes virginicus(Ulyshen and Shelton, 2012) | ||||
| Attractant | Trogoderma granariumCohen et al., 1974) | ||||
| Major metabolic component | Termites(general; Macháček et al., 2023) |
Volatile lactones and phenolic compounds are important in the early discovery of baits. For example, 6-pentyl-2H-pyran-2-one and its derivatives have been shown to attract thrips (El-Sayed et al., 2014) and cockroaches (Szőri et al., 2008). Similarly, phenylpropanoids, such as trans-isoeugenol and related eugenol derivatives, are well-known attractants of rootworms (Whitworth et al., 2002), fruit flies (Royer et al., 2018), and subterranean termites (C. curvignathus; Achmad et al., 2021). However, phenolic compounds have toxic effects on wood-destroying organisms in a dose-dependent (Putri et al., 2025; Zalsabila et al., 2024). High concentrations of eugenol: Clove leaf extract containing 7.92% of eugenol actually slowed termite response (Indrayani et al., 2018) and may contribute to the anti-termite activity of Azadirachta excelsa seed kernel extract (Adfa et al., 2023). This suggests that the volatile lactones and phenolics in rubberwood potentially act as low-dose attractants, providing directional cues that facilitate the interception of bait by foragers.
Once the bait has been located, the fatty acids perform multiple functions. Arachidonic acid, a polyunsaturated fatty acid, is an essential precursor of eicosanoids that regulate insect development and immunity (Vatanparast et al., 2018). Its presence in rubberwood likely provides a nutritional phagostimulant cue that encourages termites to continue feeding. Methyl palmitate has dual ecological roles as both an attractant (Le Conte et al., 1989) and a kairomone (Tebayashi et al., 2003). At the same time, it has been reported to possess antibacterial properties (Arinana et al., 2025), acaricidal activity against mites (Wang et al., 2009, 2010), and growth-inhibitory effects on insects (McFarlane and Henneberry, 1965). Termites may have attractive and nutritional roles, similar to other fatty acids that promote probing and feeding (Cohen et al., 1974). Methyl oleate has been linked to colony recognition and communication in Macrotermes gilvus (Kuswanto et al., 2015), suggesting that fatty acid signals in rubberwood help regulate non-agonistic interactions in the bait.
In termites such as Coptotermes formosanus, oleic acid is widely recognized as a necromone that triggers corpse burial behavior mediated by odorant-binding proteins (Li et al., 2024). However, their ecological functions are not restricted to avoidance. In Reticulitermes virginicus, oleic acid has been shown to stimulate building and covering behaviors, leading to the deposition of soil sheeting over treated substrates (Ulyshen and Shelton, 2012). In our field tests, rubberwood stakes were consistently covered by extensive soil sheeting compared with jabon or acacia [Fig. 4(a–c)]. Soil sheeting protects foragers against predators and desiccation, thereby creating a favorable microenvironment. Oleic acid also acts as a general attractant in stored-product beetles (Cohen et al., 1974) and is a major metabolic component of termite lipids (Macháček et al., 2023). These multiple roles explain why rubberwood baits attract termites; oleic acid may influence termite–bait interactions through hygienic, constructional, or substrate-modifying behaviors that could indirectly affect bait persistence and accessibility under field conditions.
Collectively, these findings indicate that rubberwood’s semiochemicals act in a synergistic manner:
-
Volatile lactones and phenols potentially guide foragers to the bait during the search phase.
-
Nutritional fatty acids potentially stimulate feeding and recruit additional workers.
Rather than being a single active metabolite, it is a blend of volatiles and fatty acids that may contribute to termite orientation and feeding behavior. Although field attack rates are also strongly influenced by physical wood properties, these chemical cues may play complementary roles in shaping termite–wood interactions (Etcheverry et al., 2025). This explains why rubberwood baits are discovered faster and consumed more heavily than other woods. This chemical complexity supports the key ecological steps of baiting: interception, mass recruitment, sustained feeding, and the colony-wide transfer of toxicants. However, further studies on the ecological role of each compound need to be conducted experimentally.
4. CONCLUSIONS
This study demonstrates that rubberwood (H. brasiliensis) has a strong potential as a bait matrix for multigenera subterranean termites in tropical regions. The higher cumulative consumption of rubberwood over time, therefore, suggests that its advantage emerges primarily during the sustained feeding phase. Importantly, the advantage of rubberwood as a bait matrix appears to emerge primarily during sustained feeding rather than during initial discovery, which is critical for effective colony-level baiting. Sustained feeding was closely linked to physico-chemical properties, including lignin–hemicellulose complexes, hardness, and extractive content. GC-MS analysis revealed a synergistic suite of semiochemicals: lactones and phenolics that potentially act as attractants and fatty acids that potentially function as feeding stimulants. Collectively, these traits explain the ability of rubberwood to accelerate interception, sustain feeding, and support colony-wide recruitment. By providing both nutritional and semiochemical advantages, rubberwood represents a promising substrate for baiting strategies that target multigenera subterranean termites, potentially reducing the duration and improving the effectiveness of termite control programs in the tropics. However, in tropical ecosystems where fungus-growing termites constitute a substantial proportion of the community, baiting outcomes may be more variable due to their distinct foraging and nutritional ecology.