
1. INTRODUCTION
The demand for solid wood from natural forests or plantation forests is continuously increasing along with population growth in Indonesia. The increasing trend in roundwood production is still not able to meet the need for roundwood raw materials. This situation is causing plantations and communities to actively search for alternative fast-growing tree species (Pertiwi et al., 2017). As an alternative, meeting the need for wood raw materials can be performed by switching to cultivated wood of fast-growing tree species. Some fast-growing tree species have been developed in Indonesia, which includes Acacia spp., Eucalyptus spp., Falcataria moluccana, Gmelina arborea, etc (Hidayati et al., 2019). Neolamarckia cadamba (Roxb.) Bosser is one of fast-growing trees with a local name of jabon putih or white jabon (Huda et al., 2019; Krisnawati et al., 2011). White jabon is widely developed because it has several advantages, including fast growth period and good wood quality (Huda et al., 2019). The wood is suitable for multiple ends uses, such as plywood, flooring, beams and rafters, boxes and crates, tea-chests, packing cases, shuttering, ceiling boards, toys, wooden shoes, bobbins, yokes, carvings, matches, chopsticks and pencils (Soerianegara and Lemmens, 1993). Studies about wood properties of white jabon have also been conducted by previous researchers (Arisandi et al., 2024; Chaerani et al., 2019; Herawati et al., 2024; Pertiwi et al., 2017). Additionally, the properties of wood modification of jabon have been conducted by previous studies (Arsyad et al., 2019; Dirna et al., 2020; Lestari et al., 2018; Priadi et al., 2020, 2021).
Based on the potential uses of white jabon wood, a breeding program of the species has been conducted in Indonesia to increase its wood productivity. The first generation (F-1) of progeny trial has been established at Forest Research of Alas Ketu, Wonogiri District, Central Java Province, Indonesia. Based on the previous measurement of stem diameter at this progeny trial, the result showed that the stem diameter was varied. A similar result was also found in another report for white jabon (Chaerani et al., 2019). This suggested that growth rate of white jabon was different among the trees, despite being planted at the same site and silviculture treatment. In addition, region and climate have significant effect on growth of Pinus densiflora from Korea (Lee et al., 2022). Pertiwi et al. (2017) reported that stem diameter and tree height of 4-year-old white jabon trees differed among the trees in a community forest.
Lateral growth rate of wood determines the strength and usefulness because it can change proportion of fibers, axial, and radial (Zobel and Buijtenen, 1989). Pertiwi et al. (2017) reported that fiber diameter, fiber wall thickness, vessel diameter, and wood cell proportion were significantly different among the three lateral growth rates in 4-year-old white jabon planted in East Java, Indonesia. The results showed that radial position affected the variation of fiber length, vessel diameter, and wood cell proportion. Additionally, fiber length and vessel diameter were increased from pith to around 6 cm and remained constant toward the bark. This suggested that xylem maturation would start when the stem diameter reached 6 cm from the pith (Pertiwi et al., 2018).
Information regarding effect of lateral growth rate and radial position on wood cell morphologies and proportions of white jabon is still limited in Indonesia, specifically trees from breeding programs such as progeny trials. Therefore, this study aimed to investigate effect of lateral growth rate and radial position on wood cell morphology and proportion of 10-year-old white jabon from progeny trial planted in Central Java, Indonesia. The results were expected to serve as a foundation for developing the uses of white jabon wood and for optimizing the breeding process.
2. MATERIALS and METHODS
A total of nine trees of 10-year-old white jabon planted in the first-generation progeny trial site in Forest Research of Alas Ketu, Wonogiri, Central Java Province, Indonesia were used as sample trees in this study. The description of the trial site is shown in Table 1. There are two factors as treatment in this study, namely 1) lateral growth rate, and 2) radial position. For the first factor, all trees in the progeny trial site were initially measured for diameter at breast height (DBH) as the basis to determine lateral growth rate. Three categories of lateral growth rate used in this study were fast growth [diameter greater than (average + SD)], medium growth [diameter between (average) ± SD] and slow growth [less than (mean – SD)] based on Ishiguri et al. (2012; Table 2). For the second factor, core samples were taken from each of the nine trees using a 5 mm diameter increment borer (Haglöfs, Långsele, Sweden). The core samples were divided based on radial position of the stem, namely near the pith, middle, and near the bark with a size of 1 cm for slide and maceration preparations.
The observed parameters of cell morphology and proportion, including fiber diameter, fiber lumen diameter, fiber wall thickness, and vessel diameter were measured using image of the transverse section of wood. The small samples were sliced using a microtome (NS-31, Yamato, Tokyo, Japan) with a thickness of 15–20 μm for transverse sections. This section was stained with 1% safranin, dehydrated, cleared using Xylol, mounted with Entellan, and then covered with cover slip. Image of these transverse sections was captured using digital camera (DP70, Olympus, Tokyo, Japan) connected to a light microscope (BX51, Olympus). The images obtained were measured using Fiji Image-J software with a total of 50 fibers and 30 vessels.
For measuring the fiber length, the small core samples were macerated using Franklin’s solution. The separated fiber was transferred into glass slide, stained with 1% safranin, washed using alcohol, mounted using Entellan, and covered with cover slip. The samples were photographed using digital camera connected to a light microscope. The images obtained were measured using Fiji Image-J software. A total of 50 fibers were measured.
Wood cell proportion, comprising fiber, vessel, axial parenchyma, and ray, was measured using total tracing area. Transverse section was captured at 20 × magnification by a digital camera attached to a light microscope. The pale tissue is detected as axial parenchyma in the area of fibers. Subsequently, images were captured using the same digital camera. A total of three areas of each image were measured and averaged for all samples. The measurement of each area was 1 × 1 mm, using Fiji Image-J software. The value of each area of wood cell was entered in the Excel to calculate the presentation of each wood cell type.
In this study, two-way analysis of variance (ANOVA) was used to analyze the two factors, namely effect of lateral growth rate and radial position, on cell morphology and proportion. For the significant effect, Tukey’s Honestly Significant Difference (HSD) was used for further testing.
3. RESULTS and DISCUSSION
The average value of fiber length, fiber diameter, fiber lumen diameter, fiber wall thickness, and vessel diameter of white jabon in this study were 1.35 mm, 22.94 μm, 18.34 μm, and 2.34 μm, and 134.24 μm, respectively (Table 3). Another study of younger age (4-year-old) white jabon by Pertiwi et al. (2017), showed that average value of fiber length, fiber diameter, fiber wall thickness, and vessel diameter were 1.52 mm, 26.9 μm, 1.84 μm, and 156 μm, respectively. Additionally, a study by Fajriani et al. (2013) on 7-year-old reported that fiber length, fiber diameter, fiber lumen diameter, and fiber wall thickness were 1.33 mm, 23.53 μm, 18.25 μm, and 2.43 μm, respectively. Another study on older age (20-year-old) of white jabon planted in Malaysia showed that fiber length, fiber diameter, fiber lumen diameter, fiber wall thickness, and vessel diameter were 1.34 mm, 34.1 μm, 24.58 μm, 5.1 μm, and 177.17 μm respectively (Ismail et al., 1995). In this study, the result for fiber length was lower compared to 4-year-old white jabon but similar to 7-year-old and 20-year-old species (Fajriani et al., 2013; Ismail et al., 1995; Pertiwi et al., 2017). However, for fiber wall thickness, the results were similar to 7-year-old and higher than 4-year-old, but lower compared to 20-year-old. This result suggests that cell wall thickness of white jabon tends to increase with increasing age. Gujar et al. (2015) reported that fiber wall thickness tend to increase from younger to older tree age on five different ages of white jabon. Fiber wall thickness of Himalayan birch was increase from 10-year-old to 25-year-old (Uyup et al., 2023). Moreover, fiber length does not always rise with the increasing age. In hardwood and softwood, differences in fiber length can be influenced by the age of the plant, the environment (nutrient and water absorption), the location of wood in the stem, and climate (Panshin and de Zeeuw, 1980; Shmulsky and Jones, 2011).
Table 3 shows the average value of cell proportion of white jabon. The average value of fiber, vessel, axial parenchyma, and ray proportion were 57.87%, 20.41%, 1.67%, and 20.05%, respectively. Pertiwi et al. (2018) reported that the average values of fiber, vessel, axial parenchyma, and ray proportion were 53%, 21%, 15%, and 11%, respectively for 4-year-old white jabon. The average value of fiber, vessel, and ray proportion were 71.83%, 16.9%, and 11.48%, respectively for 20-year-old white jabon from Malaysia (Ismail et al., 1995). In this study, fiber proportion was lower compared to the report by Ismail et al. (1995), but higher than the younger age reported by Pertiwi et al. (2018). In the case of proportion of ray cell, the result in present study was higher compared to the previous report by Ismail et al. (1995). This shows that fiber proportion of white jabon increased with increasing age. However, the ray and vessel proportion decreased with age. Ray and vessel frequency of white jabon tended to decrease from five different age from younger to older (Gujar et al., 2015). It suggesting that decreasing of ray and vessel frequency also affected decreasing on ray and vessel proportion, while increasing of fiber proportion.
The significant differences among three categories of lateral growth rate and radial position of white jabon are shown in Table 3. Lateral growth rate has a significant effect on fiber diameter and fiber lumen diameter, as well as vessel diameter and vessel proportion. However, radial position has significant effect on fiber length, fiber diameter, and fiber wall thickness, as well as vessel diameter and proportion.
Pertiwi et al. (2017) reported that fiber wall thickness, vessel diameter, fiber proportion, vessel proportion, ray proportion, and axial parenchyma proportion of white jabon were significantly different for the three lateral growth rates. In this study, similar result was found for vessel diameter and vessel proportion. A comparison of cell morphology and cell proportion is shown in Figs. 1–9. Fig. 1 shows the value of fiber length in three categories of lateral growth rate and radial position. Based on the results, no significant difference in fiber length among three categories was found. This suggested that fiber length was independent of lateral growth rate. Pertiwi et al. (2017) reported that lateral growth rate factor had no significant effect on fiber length. Additionally, wood fiber elongation in white jabon is not affected by lateral growth rate. Rapid growth on hardwoods does not consistently produce shorter fibers when the comparisons are made based on wood of the same age (Zobel and van Buijtenen, 1989). Fiber diameter of the fast growth is significantly higher when compared with the slow and medium growth (Fig. 2). Fiber lumen diameter is significantly higher compared to the slow and medium growth (Fig. 3). These results are in line with the study reported by Roque and Fo (2007), where there was a significant influence of growth rate factor on the fiber lumen diameter in G. arborea planted in Costa Rica. Lateral growth rate did not affect fiber wall thickness (Fig. 4). Vessel diameter of the slow growth was significantly different from the medium and fast growth (Fig. 5). Pertiwi et al. (2017) reported that there was a significant influence between lateral growth rate factors on vessel diameter. This can be influenced by the differences in age and location where the trees grow, despite being the same species. Fiber, axial parenchyma, and ray proportion did not differ among three growth categories (Figs. 6, 8, and 9). Vessel proportion of slow growth was significantly different from the medium and fast growth (Fig. 7). This result suggests that increasing lateral growth rate will affect the higher fiber diameter, fiber lumen diameter, vessel diameter, and vessel proportion.
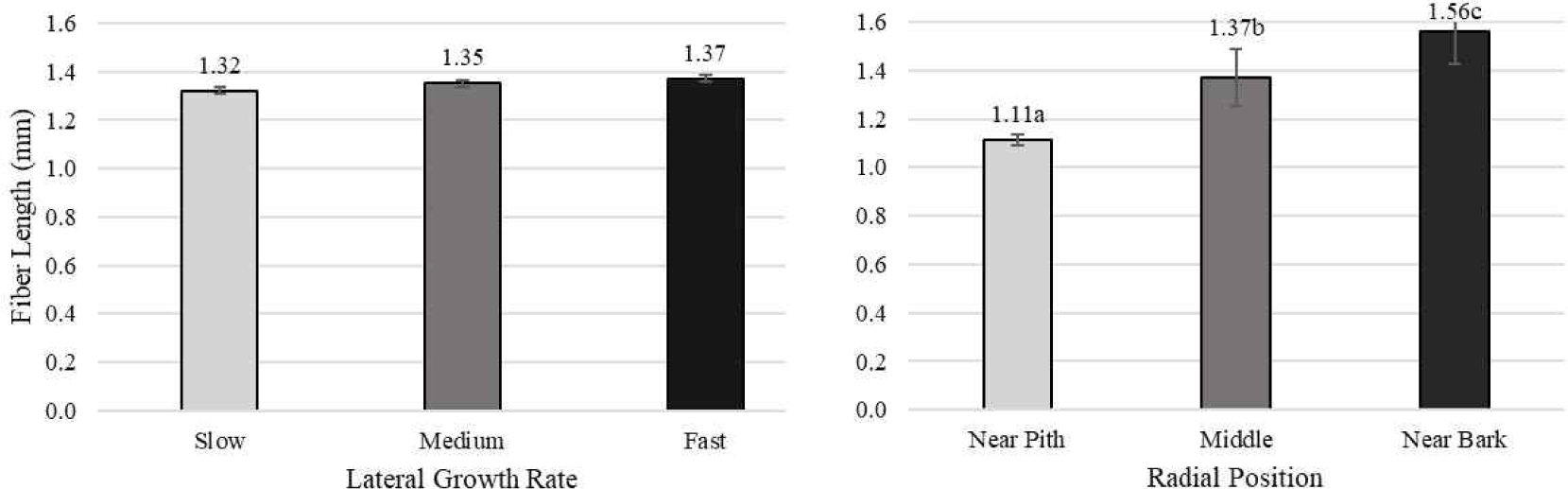
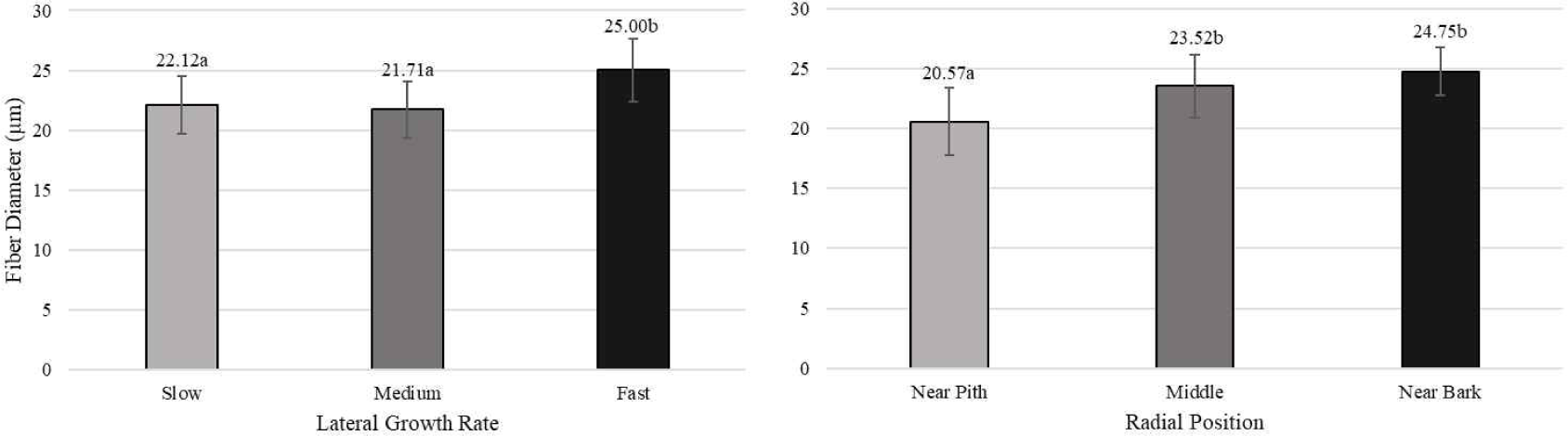
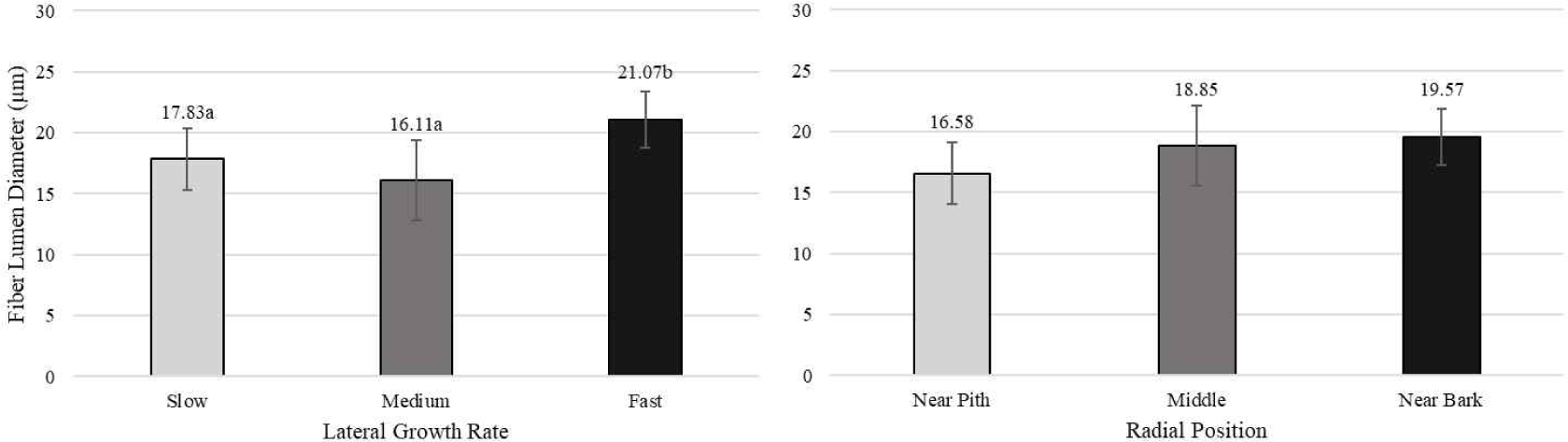
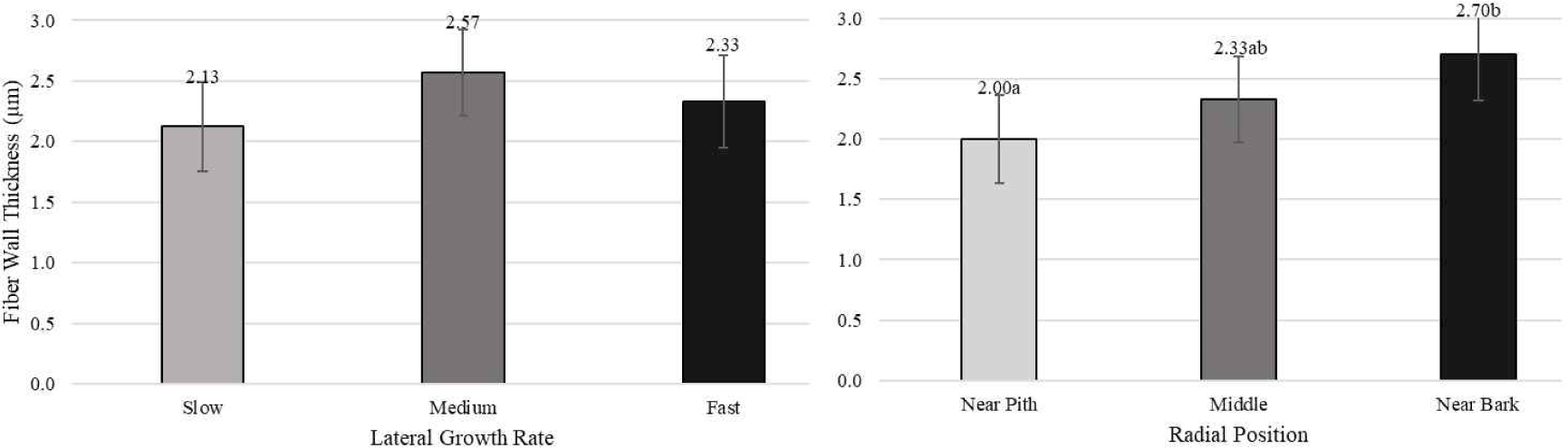
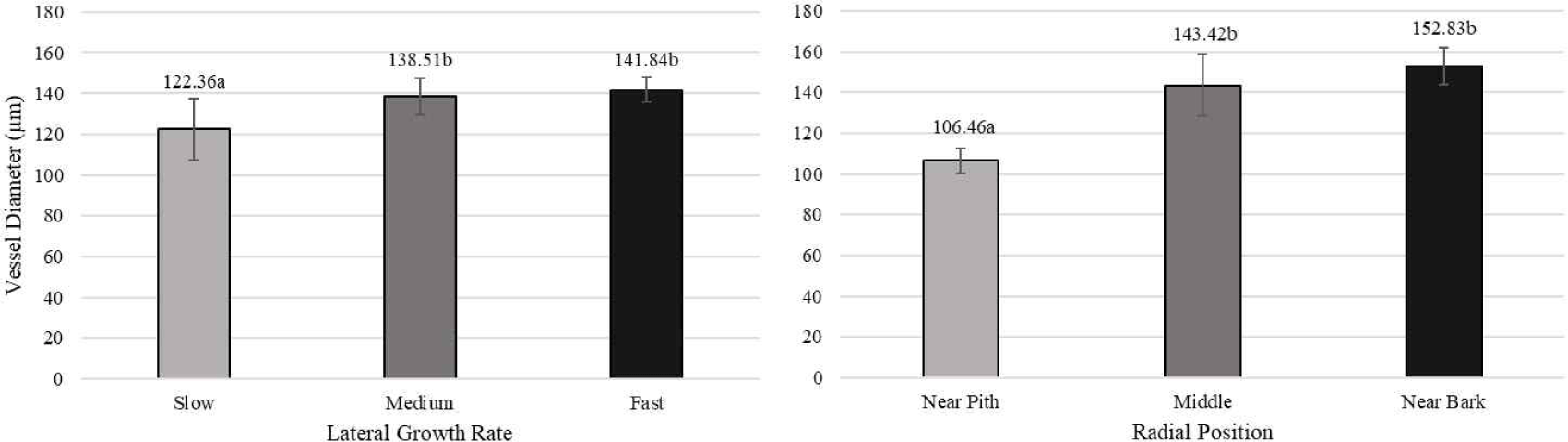
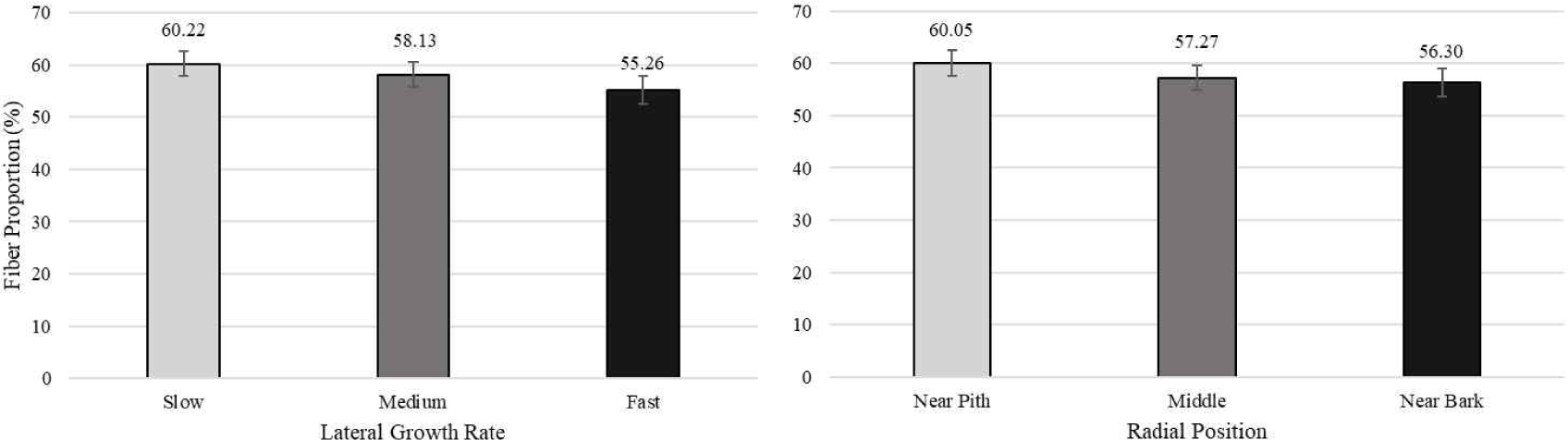
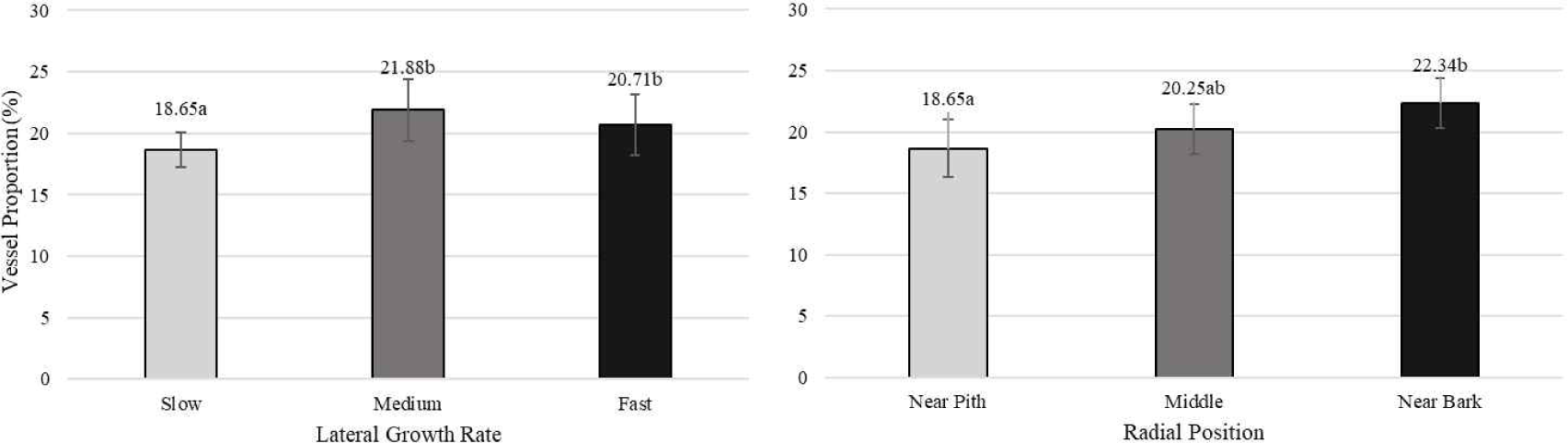
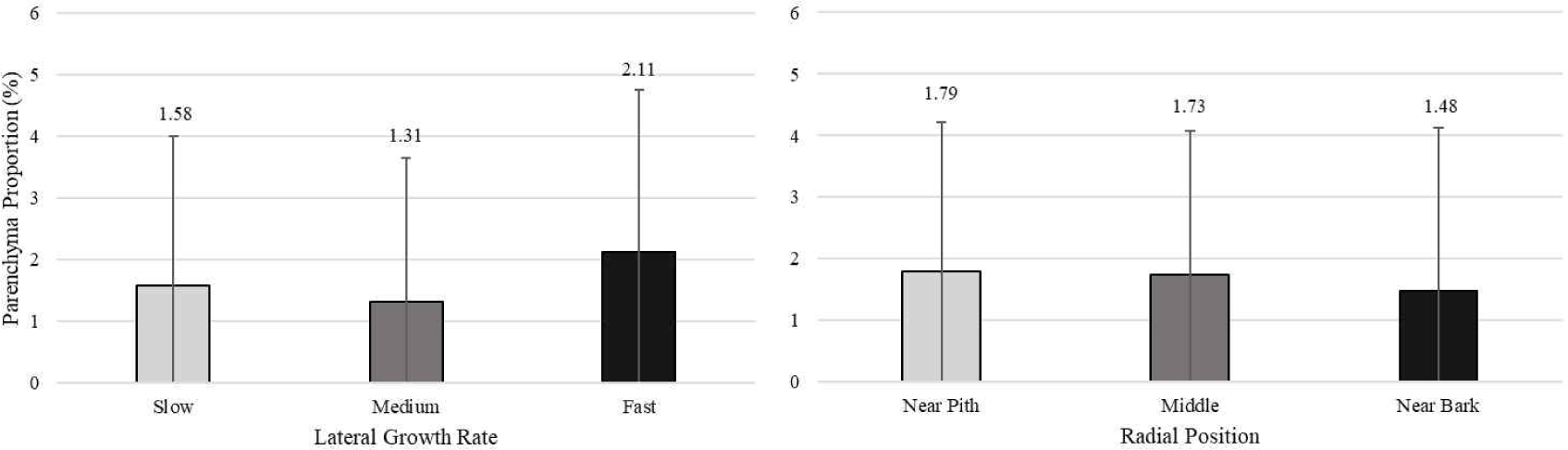
Radial position has a significant effect on fiber length and fiber diameter, including vessel, ray, and fiber proportion of white jabon (Ismail et al., 1995). Fiber length, fiber diameter, fiber lumen diameter, and vessel diameter tended to increase from pith to bark (Figs. 1–4). Pertiwi et al. (2017) reported that fiber length of white jabon rapidly increased by approximately 4–6 cm from pith. In this present study, fiber length increased significantly from pith to bark (Fig. 1). A similar trend of the fiber length was found in 20-year-old white jabon (Ismail et al., 1995). Fiber diameter and vessel diameter increased from pith until middle part and remained constant in the bark (Figs. 2 and 5). For the fiber wall thickness, it increased significantly from pith to bark (Fig. 4). Similarly, Pertiwi et al. (2017) reported that fiber wall thickness increases from pith to bark, accompanied by the xylem maturation process beyond 6 cm from the pith. Regarding fiber, axial parenchyma, and ray proportion, the parameters were almost constant from pith to bark (Figs. 6, 8, and 9). Vessel proportion increased from pith to bark (Fig. 7). A similar result of radial variation of fiber, axial parenchyma, and ray proportion was found by Pertiwi et al. (2017). Several of cell morphology of white jabon increased from pith to middle and remained constant in the bark. This result was in line with the previous report by Pertiwi et al. (2018). The results on several cell morphologies increased from pith to approximately 4–6 cm distance and tended to remain constant until the bark. This suggests that the formation of mature wood of white jabon starts in 4–6 cm from the pith (Pertiwi et al., 2018). The increase in cell morphologies from pith to middle which remains constant to the bark in this study, suggests the formation of mature white jabon. Among all the measured characteristics, no interaction between lateral growth rate and radial position was found, showing that the two factors did not influence each other.
The results of this study showed that lateral growth rate and radial position affected some wood cell morphology and proportion of the 10-year-old age of white jabon. However, fiber length, fiber wall thickness, and fiber proportion were not significantly different among the three lateral growth rates. This suggested that the increasing tree growth of white jabon did not often decrease the quality of wood anatomical properties. Therefore, tree growth can be developed by applying silviculture treatment or using genetically improved seeds from the breeding process on the plantation. Effective genetic selection during the breeding process can also be practiced to improve wood quality.
4. CONCLUSIONS
This study investigated effect of lateral growth rate and radial position on wood cell morphology and proportion of nine white jabon trees of 10-year-old N. cadamba planted in progeny trial in Central Java, Indonesia. The results showed that lateral growth rate had a significant effect on fiber diameter, fiber lumen diameter, vessel diameter, and vessel proportion. Among the three categories (slow, medium, and fast growth), the medium and fast growth showed better anatomical characteristics. Based on these results, faster growth rate did not often decrease wood quality (anatomical characteristics) in white jabon. Radial position had a significant influence on fiber length, fiber diameter, fiber wall thickness, vessel diameter, and vessel proportion. The increased cell morphologies from pith to middle which remained constant to the bark, suggested the formation of the mature wood of white jabon. Additionally, the interaction between lateral growth rate and radial position did not make a significant difference to morphology and proportion of white jabon wood cell.