
1. INTRODUCTION
Larix kaempferi (Lamb.) Carrière was first introduced to Korea in 1904 for reforestation and wood utilization (National Institute of Forest Science, 2019). However, the domestic forests were greatly damaged during the Japanese colonial period and the Korean War and have been devastated significantly due to the abusive logging and frequent disasters. The first 10-year forest reforestation plan was established in 1973 for the reforestation of destroyed forests (Korea Forest Service, 2013). Since 1975, afforestation has been carried out to secure a supply of fuel for forestry in rural areas and to restore devastated mountain areas, by designating March 21-April 20 every year as the national planting period (National Archives of Korea, 1974). Efforts have also been made to protect forests by strongly controlling illegal logging. The 10-year forest reforestation plan, which initially had been planned until 1983, was successfully achieved in 1978 at the reforestation of 108 ha, which was more than the reforestation goal of 100 ha, four years earlier than the plan. At that time, the government planted fast growing trees: Populus tomentiglandulosa and Populus euramericana, and trees for wood use: Pinus koraiensis, Cryptomeria japonica, Chamaecyparis obtusa, and Larix kaempferi, etc. (Korea Forest Service, 2013). Larix kaempferi is still recommended as a planting species for economic forestry and it is a species planted nationwide except for the southern coastal region (National Institute of Forest Science, 2019). According to a 2020 survey, the area of Larix kaempferi trees planted in Korea by 2019 was 4,559 ha, which is the second most planted species following cypress (4.952 ha) (Korea Forest Service, 2020).
Larix kaempferi has a high value as a commercial purpose and is the second largest supply following pine tree. It has excellent water resistance and durability, and the price of lumber is relatively cheap, so it is widely used for building materials, ships, raw wood, plywood agricultural tools, pulp, and pellets (Kim et al., 2015; Lee et al., 2018; National Institute of Forest Science, 2019). In the case of pellets, Larix kaempferi is also a representative tree species that can be continuously supplied if it is used as a raw material for them (Kang et al., 2018; Yang et al., 2018a, 2018b). Recently, studies related to drying, heat treatment, and bending performance, etc. have been reported to increase the utilization of Larix kaempferi wood as a building material (Kim et al., 2018; Park et al., 2018; Lee, 2020; Park et al., 2020).
The suitable planting period for Larix kaempferi recommended by Korea Forest Service is between March 11 and April 30 in considering physiological characteristics and seasonal conditions (Korea Forest Service, 2000). This is because the condition right before the bud as the frozen ground during the winter melts is a suitable condition for planting. On the other hand, the recommended optimal felling period is from November in the current year to February in the following year when the growth stops. This is because, if felling is carried out in spring or summer when the moisture content in the wood is high, it is easily damaged by fungi or pests, so that the value of the wood decreases and drying occurs easily (Viitanen, 1997; Rydell et al., 2005; Brischke and Rapp, 2008). After all, the growth period of trees provides important basic data for determining the planting and felling time. The initiation and termination of radial growth of trees are determined by cambial activity, and the timing varies depending on the tree species and growing region (Deslauriers et al., 2003; Gričar et al., 2005; Rossi et al., 2006; Seo et al., 2007; Liang et al., 2009; Oberhuber and Gruber, 2010; Rathgeber et al., 2011; Seo et al., 2011). For this reason, many studies on monitoring of the period of cambial activity, which is important data to decide the timing of reforestation and felling, by tree species and region were fulfilled, however, the study on the domestic studies is not sufficint (Park and Seo, 2000; Kwon and Kim, 2005; Park et al., 2015; Seo et al., 2017). The study on the cambial activity of Larix kaempferi, which is a major planted species in Korea, has been published only one time so far (Park and Seo, 2000).
The radial growth of trees occurs when secondary xylem and secondary phloem are formed by proliferation of cambial initial cells, xylem mother cells, and phloem mother cells constituting the cambial zone (hereafter, cambium) (Bhat et al., 2001; Plomion et al., 2001; Wodzicki, 2001). Secondary xylem is formed through the enlargement, thickness, and lignification processes of the cells divided from cambial initial cells and xylem mother cells (Wilson, 1970; Denne and Dodd, 1981; Malik, 2020). The timing of precipitation is an important factor influencing the initiation and termination of cambial activity in tropical or subtropical regions with distinct dry and raining seasons, (Bullock, 1997; Borchert, 1999; Dié et al., 2012), but the temperature is known as an important factor inducing the initiation of cambial activity in the temperate and cold regions with four distinct seasons (Deslauriers et al., 2008; Gričar et al., 2007; Rossi et al., 2006; Seo et al., 2007; Thibeault-martel et al., 2008). In some studies, the effect of temperature on the initiation of cambial activity was investigated using the heat summation (degree-days or heat sum) rather than the daily temperature condition (Sarvas, 1972; Seo et al., 2008; Seo et al., 2017).
Domestically, the systemic research on the monitoring of cambial activity was started with Larix kaempferi, Pinus rigida, Pinus densiflora in the Mt. Worak in Chungcheongbuk-do (Park and Seo, 2000). After that, the monitoring results of cambial activity for Pinus densiflora, Pinus koraiensis, Quercus mongolica, Robinia pseudoacacia, Cornus controversa, and Prunus sargentii in Chuncheon area were compared each other, depending on tree species (Kwon and Kim, 2005). The cambial activity of Pinus koraiensis and Chamaecyparis pisifera in Mt. Worak was analyzed as well (Seo et al., 2017). A comparative study on the period of cambial activity depending on regions was also conducted using Pinus densiflora in Anmyeon-do, Taean-gun, and Sogwang-ri, Uljin-gun (Park et al., 2015). Recently, the cambial activity of the major conifers (Abies koreana, Pinus koraiensis, Taxus cuspidata, and Picea jezoensis) distributed in the sub-alpine zone of Deogyusan National Park was monitored and the results were compared based on the tree species (Park et al., 2021). However, studies on the same species in different regions on the cambial activity have been insufficiently compared.
The present study was conducted to monitor the cambial activity of Larix kaempferi growing in different regions in Chungcheongbuk-do (hereafter Chungbuk) to establish basic data necessary to determine the optimal planting and felling period for each region. Also, the relationship between temperature and the initiation of cambial activity was investigated to elucidate how the predicted temperature change in the future can affect the change in the growth initiation of Larix kaempferi.
2. MATERIALS and METHODS
Larix kaempferi forest units planted in Chungbuk National University Academic Forest (128°05′50″E, 36°50′12″N) (Mt. Worak; hereafter WRLK) Deoksan-myeon, Jecheon-si, Chungcheongbuk-do, and Chungcheongbuk-do Forest Environment Research Institute (178°40′21″E, 36°37′34″N) (Mt. Midong; hereafter MDLK) located in Miwon-myeon, Cheongju-si, Chungcheongbuk-do were selected as research sites (Table 1).
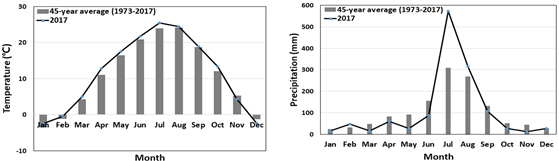
According to the meteorological data from the Jecheon meteorological station closest to the Mt. Worak research site, the average annual temperature for the last 45 years (1973-2017) was 10.2℃, and the annual precipitation was 1335.2 mm (Fig. 1). The temperatures of the coldest month (January) and the warmest month (August) were -5.2°C and 23.8°C, respectively.

The meteorological data from Boeun Meteorological Center closest to the Mt. Midong research site reported that the average annual temperature for the last 45 years (1973-2017) was 10.9℃, and the annual precipitation was 1276.2 mm (Fig. 2). Similar to data from the Jecheon meteorological station, the coldest month and the warmest month were January (-3.5℃) and August (24.1℃), respectively. The precipitation was the lowest in January with 25.1 mm and the highest in July with 308.9 mm.
Five dominant tree in Larix kaempferi stand at each research site were selected for cambium monitoring. The method, generally used in the cambium monitoring research, was applied to this study (Rossi et al., 2006; Gruber et al., 2009; Park et al., 2021). The average diameter at breast height of the Larix kaempferi at Mt. Worak was 35.2 (±7.3) cm, which was smaller than the average diameter at Mt. Midong, but the average tree height at Mt. Worak was 22.4 (±1.8) m, slightly higher than the average tree height of 21.4 (±0.9) m at Mt. Midong (Table 1).
Heat summation was utilized to analyze the role of temperature in inducing the initiation of cambial activity. The heat summation was calculated by accumulating the temperature on days when the average daily temperature was higher than or equal to 5℃, which tree growth is possible, as shown below (Sarvas, 1972; Seo et al., 2008; Seo et al., 2017). In the case of Mt. Worak, there was no available data on the temperature measured at the research site, so data from the Jecheon meteorological site were used, and for Mt. Midong, temperature data measured on site (HOBO) were used.
Where 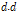 is cumulated value of
is cumulated value of 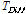 ,
, 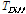 is the difference between the average temperature of the
is the difference between the average temperature of the 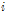 th Julian day and +5°C,
th Julian day and +5°C, 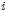 is the Julian day on which the average daily temperature first became to be higher than +5°C and lasted for 5 days or more in that condition.
is the Julian day on which the average daily temperature first became to be higher than +5°C and lasted for 5 days or more in that condition.
Samples were collected using a mini trephor with 1.9 mm diameter, so that the cambium and at least two recent tree rings of the secondary xylem were included (Fig. 3(A), 3(B)). Sample collection was started on April 7, 2017, at Mt. Worak and Mt. Midong, and the cambium samples was collected at weekly intervals until the day that collection ended. The last sampling dates were October 6 at Mt. Worak, and September 29 at Mt. Midong. The collected samples were stored in distilled water at the same time as they were extracted from the trees, and refrigerated until embedding to prevent drying of the samples.

The transverse thin sections of the small specimen of cambium were prepared to observe the chronological cambial activities and the secondary xylem development by the season. Before embedding, the cross-sections of the small specimens were labeled with a pencil to facilitate the observation of the cross-section (Fig. 4(A)). Then, after the cambium small specimens were immersed in a mixture of distilled water and polyethylene glycol (PEG2000) at 1:3 ratio (Fig. 4(B)), they were placed in a thermo-hygrostat (OF-12G, Jeiotech, Korea) set at 60°C for 2 to 3 days, so that PEG2000 slowly penetrated into the cell lumen of the small specimen as distilled water evaporated (Fig. 4(C)). The small specimens of cambium prepared as above procedures were fixed in the embedding frame using 100% PEG2000 (Fig. 4(D)). When fixing, the small specimen pieces were placed for their cross-section to be oriented vertically.

The thin slices of the embedded samples were prepared using a sliding microtome (H/I sliding microtome, Hacker, U.S.A) with a thickness of 10-15 µm (Fig. 5(A)). The thin sections were stained with a mixture of safranine (1%) and astra blue (0.5%) for about 1 minute to help the observation of the cambium and the secondary xylem development. Safranine stains all xylem cell walls in red, while astra blue stains the xylem cell wall without lignin deposition in blue, which helps to observe the degree of cell development (Gärtner and Schweingruber, 2013). The stained sections were washed and sealed with cover glasses using glycerin (Fig. 5(B)).

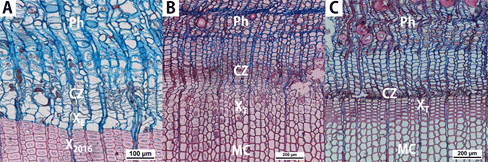
An optical microscope (Eclipse 80i, Nikon, Japan) connected with a camera (Nikon, TH/D5500 15-55VR Ⅱ KIT) was used to observe the wood cells. In this study, the initiation period for cambial activity was defined as when the enlargement of the secondary xylem cells (XE in Fig. 6(A)) divided from cambial zone (CZ in Fig. 6) is exhibited. The termination of cambial activity was defined as when no further secondary xylem cell division occurred in the cambium (Fig. 6(C)).
3. RESULTS and DISCUSSION
Accurate monitoring of cambial activity was impossible in one specimen of Larix kaempferi from each site, Mt. Worak (WRLK05) and Mt. Midong (MDLK05), because a traumatic resin canal was observed. For this reason, each of the 4 specimens in which the normal cambial activity and the secondary xylem cell development process were confirmed, was used in the further analysis of the results.
Two out of four specimens of Larix kaempferi from Mt. Worak (WRLK 02 and WRLK 03) and 3 out of 4 specimens from Mt. Midong (MDLK 02, MDLK 03, and MDLK 04) showed the initiation of cambial activity on April 28th. The remaining 2 Larix kaempferi samples from Mt. Worak (WRLK 01, WRLK 04) and the remaining 1 sample from Mt. Midong (MDLK 01) showed the initiation on May 4, a week later than the others. Therefore, it was observed that the cambial activity of Larix kaempferi at Mt. Worak and Mt. Midong started between late April – early May (Fig. 7). Accumulated temperature till April 28 at Mt. Worak and Mt. Midong was 196.4 and 204.7, respectively, and heat summation till May 4 was 271.8 and 277.3, respectively, showing no significant difference (Fig. 7).
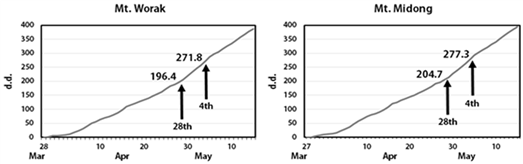
According to the previous monitoring results of cambial activity surveyed at Mt. Worak, the initiation of cambial activity of Larix kaempferi, Pinus densiflora, and Pinus rigida was confirmed to be in mid April, late April, and early May, respectively (Park and Seo, 2000), and that of Pinus koraiensis and Chamaecyparis pisifera was confirmed to be in mid April, and late April (Seo et al., 2017). It was confirmed that the initiation period of Larix kaempferi determined by the current study was similar to Pinus densiflora, Pinus rigida, and Chamaecyparis pisifera, and later than the results for Larix kaempferi and Pinus koraiensis in the previous study. Because the climate is different each year, the comparison of the dates of cambial activity initiation investigated in different years does not reflect the climatic conditions of the current year. However, because an important influencing factor to induce the initiation of cambial activity is the temperature (Deslauriers et al., 2008; Gričar et al., 2007; Rossi et al., 2006; Seo et al., 2007; Thibeault-martel et al., 2008), the utilization of accumulated temperature could allow for the comparison with other species in the same area. Among the studies conducted at Mt. Worak, there have been cases in which the accumulated temperature was applied to the investigation of the initiation of cambial activity in Pinus koraiensis and Chamaecyparis pisifera (Seo et al., 2017). According to this study, the initiation of cambial activity of Pinus koraiensis and Chamaecyparis pisifera was possible when the accumulated temperature was 134 and 200, respectively. The initiation of cambial activity of Larix kaempferi of Mt. Worak determined in this study was confirmed to be possible when the accumulated temperature was 196.4, which is similar to that of Chamaecyparis pisifera. In the case of Larix kaempferi at Mt. Midong, there was no study conducted in the same area, so comparative analysis was not possible. However, since the results were confirmed to be similar to those of Larix kaempferi at Mt. Worak, it is expected that the confirmed results can be applied in the same way.
Unlike Mt. Midong, the weather station used to calculate the accumulated temperature of Mt. Worak is about 30 km away from the site, and a slight difference from the temperature of the site should be expected. Temperature measurements for accurate calculation of accumulated temperature in Mt. Worak area should be considered in future research.
The termination of cambial activity for Larix kaempferi at Mt. Worak was started on August 4 (WRLK 01 and WRLK 03) and observed on August 18 (WRLK 04) and 25 (WRLK 02) (Fig. 8). The termination of cambial activity was confirmed on August 4 (MDLK 04) for Larix kaempferi at Mt. Midong as well, but unlike Mt. Worak, the cambial activity was terminated one by one at weekly intervals, and it was confirmed that the cambium activity was completely terminated by September 1 (MDLK 01). Therefore, the termination period of cambial activity was found to be somewhat later in Larix kaempferi at Mt. Midong compared to Mt. Worak. As a result of calculating the difference between results in this study and previous investigation on the initiation period of cambial activity, the cambium activity period of the Larix kaempferi of Mt. Worak was 104.75 days (93-120 days), which was 5 days shorter than 111.25 days (99-121 days) of Larix kaempferi of Mt. Midong.
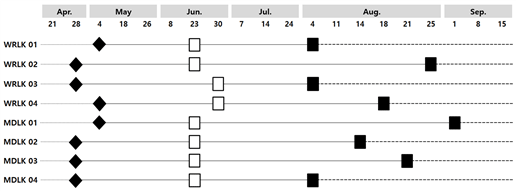
The terminations of cambial activity of Pinus koraiensis and Chamaecyparis pisifera at Mt. Worak occurred from September 15 to 22 and from August 18 to September 22, respectively (Seo et al., 2017). This is 2 weeks later for Chamaecyparis pisifera and 6 weeks later for Pinus koraiensis than on August 4, which was confirmed as the earliest day of the termination of cambial activity in the present study. For the period of cambial activity, that of Larix kaempferi was confirmed as more then 40 days shorter than Pinus koraiensis (157 days) and Chamaecyparis pisifera (145–147 days) (Seo et al,. 2017). Thus, the cambial activity period for Larix kaempferi was significantly shorter than those of Pinus koraiensis and Chamaecyparis pisifera, and the main reason was considered that its termination of cambial activity occurred earlier than other species. This was investigated to be similar to Mt. Midong, which is about 50km away from Mt. Worak.
It was reported that the optimal planting time taking into account of the physiological activity of trees is just before the bud break (Korea Forest Service, 2000, 2013). According to a previous study on Pinus sylvestris, its cambial activity was found to occur 2-5 weeks after the bud break (Seo et al., 2011). If the time of the bud break for Larix kaempferi at Mt. Worak and Mt. Midong would be estimated based on the results of this previous study, it can be predicted to be after late March to mid April, about 2-5 weeks earlier than April 28, when its cambial activity was confirmed. Therefore, it was considered that the appropriate time for planting Larix kaempferi trees in the Chungbuk area, where this study sites are located, would be from March 11, following the recommendations of the Korea Forest Service. However, no later than end of March is desirable, when the earliest tree activity is expected.
Generally, the recommended optimal period for felling is from November in the current year to February in the following year when the growth stops. (Viitanen, 1997; Rydell et al., 2005; Brischke and Rapp, 2008). It is to minimize the moisture content in trees and damage from fungi or pests after felling, and to increase the efficiency of drying. In considering the cambial activity period of domestic conifers investigated so far, it was confirmed that the already known felling period was well applied to conifers. However, unlike other conifers, Larix kaempferi in Mt. Worak and Mt. Midong areas were found to complete all cambial activity in early September, so it was reasoned that the optimal period for felling is from October.
4. CONCLUSION
It was confirmed that the cambial activity of Larix kaempferi planted in Mt. Worak and Mt. Midong started from late April to early May, and the termination of cambial activity started in early August and ended in early September. Based on these results, it is desirable to complete the planting of Larix kaempferi in the Chungbuk region before April, and it was considered that possible felling period is from October to the resting period of the following year. Through this study, the heat summation affecting the initiation of the cambial activity of Larix kaempferi in Mt. Worak and Mt. Midong was suggested, but further long term monitoring would be necessary to obtain more reliable results. Especially, an additional study will be required to find the reason that the significantly earlier termination of cambial activity in Larix kaempferi than that in other conifers (Pinus koraiensis and Chamaecyparis pisifera) confirmed in Mt. Worak is from the characteristics of the species or the from the climate conditions.
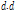 는
는 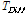 의 누적값,
의 누적값, 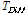 는
는 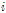 번째 율리우스 일의 평균온도와 +5℃ 차이,
번째 율리우스 일의 평균온도와 +5℃ 차이, 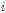 는 일평균기온이 처음으로 +5℃ 이상 5일 이상 지속된 기간이 시작되는 율리우스 일이다.
는 일평균기온이 처음으로 +5℃ 이상 5일 이상 지속된 기간이 시작되는 율리우스 일이다.