
1. INTRODUCTION
A total of 63.2% of Korea’s land is covered with forests, of which 32% are deciduous forests (Korea Forest Service, 2016). Oak trees, the dominant species with broad leaves, account for 25% of the deciduous forests. Representative tree species of oaks include Quercus mongolica, Q. dentata, Q. acutissima, Q. variabilis, Q. aliena, Q. serrata they are trees of economic value that have been widely used since ancient times to obtain forestry products (mushrooms) and make musical instruments, fuel materials, oak barrels. However, the damage to oak trees is increasing due to recent changes in climate, new types of diseases, and insect pests.
The first reported case of oak wilt disease to cause the sudden death of oak trees in Korea was published in Seongnam, Gyeonggi-do in 2004 (Son et al., 2014). Thereafter, 268,000 trees died due to oak wilt disease in 2012 and 309,000 trees died in 2013, showing an increase of 41,000 trees. Raffaelea quercus-mongolicae, the pathogen that causes oak wilt disease, infiltrates into the sapwood area of oak trees via the insect vector Platypus koryoensis. The infiltrating pathogens multiply and prevent the movement of water, causing the oak trees to desiccate to death (Kim et al., 2009; Kang and Shin, 2017). Furthermore, this disease is primarily found in Q. mongolica, Q. aliena, and Q. serrata and it is also known to often damage other oaks, such as Q. acutissima, Q.variabilis, and Q. dentata (Kim et al., 2016).
The damage caused by oak wilt disease is also occurring at a serious rate in Japan, and it has been reported that a large number of oak trees were killed by the insect vector P. quercivorus and pathogen R. quercivora (Kubono and Ito, 2002).
Diverse studies have been conducted on oak wood, including those on anatomical, physical, mechanical, and chemical properties (Kim et al., 2006; Han et al., 2015; Kim et al., 2015; Chang et al., 2017; Jeong et al., 2017). On the other hand, studies on trees damaged by oak wilt disease are somewhat limited to pest control, physical, and mechanical characteristics (Kim et al., 2011; Son et al., 2014; Kim et al, 2016; Kim et al., 2020). Although there are studies on trees infected with oak wilt disease, basic anatomical data that can facilitate the utilization of the damaged tree is insufficient. Therefore, in this study, the anatomical characteristics of damaged Q.mongolica (infected and dead) trees were compared with those of sound trees and analyzed to obtain basic data for the utilization of trees damaged by oak wilt disease.
2. MATERIALS and METHODS
To observe the anatomical characteristics of the trees affected by oak wilt disease, two Q. mongolica trees (one infected and one dead) were selected, which were confirmed to be damaged by oak wilt disease among the oak trees that grow wild in Dongtan-dong, Hwaseong-si. To compare the anatomical characteristics of infected and dead oak trees with those of sound trees, one sound tree was selected from Chuncheon-si, Gangwon-do. For the experiment, a 3-cm-thick disk was produced immediately after harvesting (Fig. 1), and the chracteristics of tree age, tree ring width, and latedwood proportion are shown in Table 1.

| Infested wood | Dead wood | Sound wood | |
|---|---|---|---|
| Tree age (year) | 59 | 55 | 58 |
| Tree ring width (mm) | 2.0 ± 1.2 | 2.0 ± 0.7 | 2.9 ± 0.8 |
| Latewood proportion (%) | 66.3 ± 20.6 | 69.2 ± 16.2 | 79.8 ± 7.8 |
The anatomical characteristics of the affected trees were observed using optical and scanning electron microscopes, and the degree of morphological infection was measured. Samples for optical microscopy were prepared by sectioning dead, infected, and sound wood from the pith to the bark at intervals of 10 annual rings. Subsequently, a slice with a thickness of approximately 15–20 μm was prepared using a sliding microtome. The prepared slices were stained with 1% safranine solution and dehydrated with an alcohol solutions (50%, 70%, 90%, 95%, and 99%). Permanent slides were then prepared and observed using an optical microscope (Carl Zeiss, DE/Axio Imager A1, Germany).
For the observation with a scanning electron microscope, clean cross-sectional samples were prepared with a microtome at 10-annual rings intervals.The prepared samples were coated with gold and then observed using a scanning electron microscope (SEMCOX EM-30, COXEM, 20kV).
To measure the length of wood fiber, a sliver of approximately 1 cm in length was prepared by separatingthe earlywood and latewood, from the pith of the bark at 5-annual intervals, which were heated and dissociated at 60°C in a mixture of hydrogen peroxide and acetic acid in a ratio of 1:1. They were washed with distilled water, dyed with methylene blue. The length of wood fibers of 30 samples of each type of wood were measured using the the DeltaPix (version 5.3.11) program, which were chosen from the annual rings of both earlywood and latewood.
Diameters of vessel elements were measured by determining tangential and radial diameters of each sample (n = 30). The tangential and radial diameters were measured by categorizing earlywood and latewood using the annual rings on a cross section at 50x magnification using the prepared permanent slides. The ratio of vessels with tylosis was calculating by estimating the number of vessels with tylosis after measuring the number of 30 vessels from each age group by dividing the wood at 10 ring intervals from the pith to the bark.
3. RESULTS and DISCUSSION
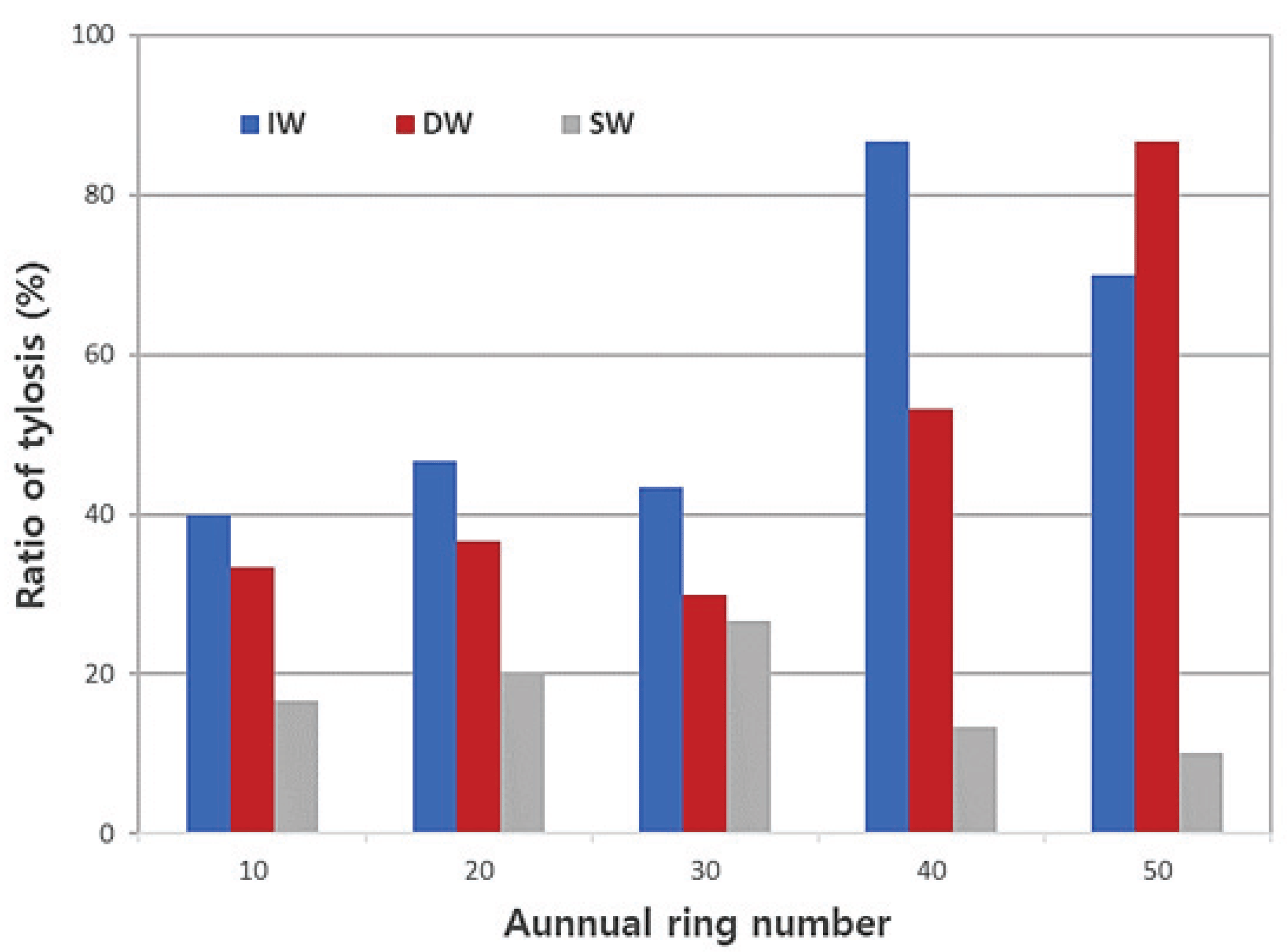
Fig. 2 shows the cross section of each annual ring of Q. mongolica infected with wilt disease, dead wood, and sound wood imaged using an optical microscope. The cross sections showed that the boundary of the annual ring was distinct due to the porous ring, and the vessels of the earlywood were large and separated and those of the latewood were small and patterned. In the infected, dead, and sound wood, tylosis was often observed in all (10th–50th) annual rings in the vessel, and tyloses in infected and dead wood showed a tendency to appear more frequently than in sound woodin the 40th and 50th annual rings. In the 10th–30th annual rings of infected, dead, and sound wood, the average ratio of tyloses were 43.3%, 33.3%, and 21.1%, respectively, and the average rates of tyloses in the 40th–50th annual rings were 78.4%, 70%, and 11.7% (Fig. 6), respectively. The results of this experiment showed trends that were consistent with optical microscope results.

Observing the cross sections using the scanning electron micrsope showed that the lumen of the vessel from the 30th and 50th rings of the infected and dead wood had mycelium (Fig. 3). Tylosis, a characteristic of oak trees, is known to block the vessel because the flow parenchyma cells adjacent to the vessel become evaginated due to an injury, while the cells are aging and transitioning into heartwood (Kwon and Kim, 2005; De Micco et al., 2017). Kuroda (2001) observed the characteristics of sapwood in the area infected with oak wilt disease and found that the distribution of Raffaella sp. and area of discolored sapwood matched, and the function of the vessel was lost. Martíneb et al. (2005) reported that mycelium was observed in the vesselin wood damaged by the pathogen. Therefore, it is believed that the oak wilt disease pathogen mainly penetrates the annual ring of the 30th or 40th sapwood portion from the bark of the tree to block the vessel and prevent the transportation of water.
Table 2 shows the length of wood fibers of Q. mongolica infected, dead, and sound wood, separated into earlywood and latewood. The average length of wood fibers in earlywood was 1.20 mm and 1.24 mm in the infected and dead wood respectively, and it was slightly longer in sound wood (1.30 mm). Furthermore, the average length of wood fiber in the latewood was similar to that of the infected and dead wood (1.28 and 1.31 mm, respectively), and was slightly longer in sound wood (1.36 mm). In all of the infected, dead, and sound wood, wood fiber length tended to increase with increasing annual rings in both earlywood and latewood, and it was found to stabilize in the 20th–30th annual rings (Fig. 4). When dividing juvenile wood and mature wood according to the variation of the length of wood fibers, it is believed to be approximately 20–30 annual rings. The wood fiber length of the tested earlywood had an average of 1.17 mm of juvenile wood and 1.31 mm of mature wood from the 25th annual ring, whereas the fiber length of the tested latewood had an average of 1.22 mm for juvenile wood and 1.39 mm for mature wood. Eom (2015) reported that the average length of wood fibers in Korean Q.mongolica was 981.4 μm (range 733– 1,323μm), which was similar to the results of this study. Meanwhile, the average length of wood fibers in Q. acutissima, Q. dealbata, Q. fenestrata, Q. lanceofolia, and Q. semiserrata were 1,199.36 μm, 1,439.37 μm, 1,479.38 μm, 1,509.75 μm, and 1,389.0 μm (Sharma et al, 2011), respectively. The length of wood fiber of Q. Suber was reported to be 1.15 mm (Sousa et al, 2009), and there was no significant difference from the length of wood fiber in this study. The IAWA Committee (1989) classified the wood fiber length of hardwood into ≤ 900 μm, 900–1,600 μm, ≥ 1,600 μm, and the wood fiber length in this study corresponds to 900–1,600 μm.
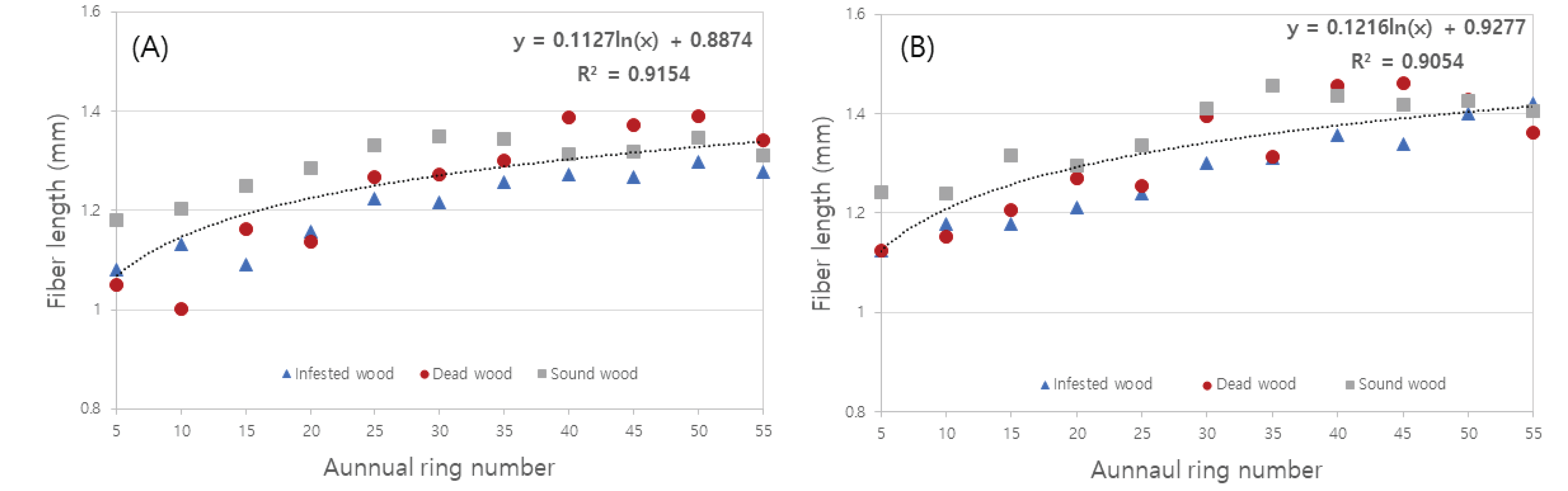
Table 2 shows the results measured by dividing the diameter of vessel elements and ratio of tylosis of infected, dead, and sound wood of oak trees into earlywood and latewood. The average tangential diameter of the vesselelement of earlywood was the smallest (234.4 μm) in the dead wood, the largest was 249.4 μm in the infected wood, the smallest radial diameter was 278.8 μm in the sound wood, and the largest was 299.6 μm in the infected wood. The average tangential diameter of the vessel element of latewood was the smallest in the sound wood at 27.7 μm and that of the dead wood was the largest at 30.5 μm; the average radial diameter was the smallest in the sound wood (35.3 μm) and that of the infected tree was the largest (38.5 μm). The radial diameters of the vesselelement in earlywood and latewood was larger than the tangential diameters. The tangential diameter of vessel elements for each annual ring of earlywood, latewood, and radial diameters was the smallest value in 10 annual rings of all of the infected, dead, and sound wood, and the diameter tended to increase as the number of rings increased (Fig. 5). Eom (2015) reported that the average diameter of the tangential line of the vessel element of domestic Q. mongolica is 224.0 μm (range = 163–286 μm), and Kim et al. (2002) reported that in sapwood of Korean Q. mongolica, the tangential diameters of vessel of earlywood and latewood were 245.2 μm and 34.0 μm, respectively, which were similar to the results of this study. Furthermore, in studies of Quercus spp., Sousa et al, (2009) reported that the tangential diameter of the vessel element of Q. suber was 133 μm, and Han et al. (2015) reported that the tangential diameters of the vessel elements of Q. rubra were 122 μm and 118 μm, respectively. The above results were smaller than the diametersof earlywood and latewood of the vessel element in this study. The IAWA Committee (1989) classified the vessel element tangential diameter as ≤ 50μm, 50–100 μm, 100–200 μm, and ≥ 200 μm, and the vessel element tangential diameter in this study was ≥200 μm. Regarding vesselelement diameter according to tree age, Lee et al. (2009) reported that both the tangential and radial diameters of vessel elements in Korean chestnut trees gradually increased with increasing tree age, and Leal et al. (2007) investigated the radial variability of the Q. suber L. vessel to report that the size of the vessel lumen increased with tree age.
The average ratio of vessel of earlywood including tylosis was the highest in infected wood at approximately 57.3% and the lowest in sound wood at approximately 17.3%. Furthermore, there was a difference in the ratio of tylosis in the order of infected, dead, and sound wood in the 10th–30th annual rings, which are considered juvenile wood areas. In the infected and dead woods, it was observed that the ratio of tylosis increased in the 40th and 50th annual rings, which are mature wood (Fig. 6). Therefore, there was a difference in the ratio of tylosis from the heartwood, which is believed to be a difference in the ability to generate tylosis between the sample trees, and it may cause death or infection.
4. CONCLUSION
This study compared and analyzed the anatomical characteristics of the damaged and dead wood affected by oak wilt disease and those of sound wood in Korea. Tylosis was frequently observed in the cross sections of infected and dead woods along with mycelium, indicating the degree of infection in the 30th–50th annual rings. There was no significant difference in wood fiber length and diameter of vessel elements among the tested woods. The wood fiber length increased to 20–30 annual rings, which qualifies as juvenile wood, and showed trends of future stabilization in the mature wood area. Infected wood had the highest average ratio of earlywood vessel including tyloses and sound wood had the lowest. In addition, infected and dead woods had the highest rate of tyloses in the 40th–50th annual rings, which was judged to be mature wood.
Therefore, there was a confirmed difference in the ratio of tylosis in the areas of juvenile and mature wood in the oak wilt-damaged wood and sound wood of Q. mongolica. It is considered that this difference in the tylosis ratio blocks water movement, causing the infection and death of the oak wilt diseased tree. In the future, it is necessary to collect samples with the exact times of infection and death to carefully observe the changes in the oak tissues.